Üks populaarne uurimisvaldkond bioloogias on see, kuidas vananemist edasi lükata või elu pikendada ja seda huvi olen näinud ka mõne bioloogiatudengi hulgas, kuid tavaliselt keskendutakse selle eesmärgiga molekulaarbioloogiale. Samas peab sellise lähenemisega arvestama kõikide eluks vajalike valkudega ning inimese ~25000 geeni moodustavad üle 100 000 valgu, kuna esialgsest RNA molekulist lõigatakse välja intronid ja ülejäänud juppide omavaheline kombineerumine annab täpselt teadmata hulgal valke, millest paljude funktsioonigi veel ei teata. Arvestades, et lisaks kuhjub vananedes hapnikukahjustusi ja väheneb üldiselt vajalike valkude hulk elusrakkudes. Kui realistlik oleks 100 000 tuhande valgu poolt läbiviidud protsesside alal hoid farmakoloogilisel korrigeerimisel? Arvestama peab inimestevahelise varieeruvusega ja kõikide nende ravimite kõrvalmõjuga.
Lisaks ei ole DNA replikatsioon täiesti täpne. Inimestel kõiguvad need hinnangud, kuid üldiselt iga DNA paljundusega muutub tuhandeid nukleotiide ja kuna ~1,5% DNAst sisaldab inimesel geene (ülejäänu on nn "junk DNA", mis võib koosneda mitteekspresseeruvates geenijääkidest, viiruste DNAst või ennast DNAs paljundavatest transponeeruvatest elementidest). Ka selles väiksemas vajalikumas osas tuleb statistiliselt keskmiselt üle 10 vea geenide peale kokku ja nendest umbes kolmandik muudab selle tulemuses valgu aminohappelist koostist. Elu jooksul võivad rakud nii palju mutatsioone koguda, et hakkavad kasvajana paljunema või muid probleemseid kahjustusi põhjustama ja selle kõige ära hoidmiseks ei paista mingit moodust. Põhimõtteliselt on võimalik, et inimeses pole isegi kahte identse DNAga rakku ja tüvirakkudelegi saab see aastakümnetega koormaks.
Ühegi ravimiga ei saa nii erinevaid muutlikke rakke normis hoida.
Elu pikendamisel peab paratamatult mõistma ka seda, mida me täpselt peame säilitama et eksistentsitunnet omada.
Teadvuse mõistmises ei paista midagi nii väga ületamatut iseenesest. Aju tekkes kasvavad eri pindmised alad ajukooreks ja teisteks aladeks, paralleelselt toimub ühenduste kasvatamine geneetiliselt programmeeritud suundade ja ajastusega. Lõpptulemusena saadakse bioloogiline protsessor mis suudab rohkemal või vähemal määral omastada infot ümbrusest, seda omavahel seostada ja vastavalt reageerida. Looduses on miljoneid liike putukatest imetajateni, kes väga erinevate närvisüsteemidega suudavad keskkonnas funktsioneerida. Iseasi kes neist teadvusel on ja mil määral, kuid läbikäivaks on see, et varieeruvusele vaatamata suudavad need kõik reaalsusega arvestada, sõltuvalt oma perifeerses ja kesknärvisüsteemi võimekusest.
Tähtsamaks küsimuseks on see, kuidas kogetu närvisüsteemis välja näeb. Holmes'i kaart annab aimduse, kuidas see nägemises kuklasagarale jõudmise ajaks muutunud on, kuid kõigi teiste meelte (lõhnad, valu, temperatuur jne.) jaoks ei ole nii mugavat ja arusaadavat enne-pärast mudelit. Ühe lahendusena paistab ajus millegi kogemisel signaalimustri ja tiheduse täpne salvestamine ja loomkatsete najal saaks juba suurema osa töö nii tehtud, kui ainult õppida sellist invasiivset skännerit looma ja ajusse sisestama liigsete kahjustusteta.
Looduses suudetakse mitmekülgsete ajudega teadvustada reaalsust ja samuti on meie ajudes loomulikku varieeruvust, mis näitab et teadvus võib küllaltki robustne olla. Üheks minimaalseks eelduseks paistab õigete juppide omavaheline ühendamine. Ühendused tajuks kasvavad umbkaudu nii, et õiged kohad saaksid õigesti ühendatud, kuid geeniekspressioonil pole nii palju täpsust aju tekkel, et iga neuronit täpselt õige rakuga ühendada. Loodud aksonikimbud samas suudavad osade endas olevate aksonitega kogemusi luua (kahtlane, et iga viimane akson oleks hädavajalik kogemuseks). Teadvuse mõistmine eeldab ühe miinimumina üldühendusteede tundmist ning tänapäeval on see lihtsam ja kättesaadavam lähenemine, sest omavahelisi ühendusi on nii närilistes, kui primaatides kaardistatud väga paljudes töödes.
Muidugi saaks teadvuse paremat mõistmist kasutada tõenäoliselt meditsiinis rohkemate ajukahjustuste raviks, kui tänapäeval.
Kui loodus saab hakkama teadvuse loomisega ja veel nii paljude eri närvisüsteemidega, siis miks inimesed kipuvad kaotama enesekindlust selle organi mõistmisel? Teiste saamatust, argust ja ajapuudust ei tohi võtta enda võimete piirina.
Kokkuvõtlikumalt ei väida ma, et teadvuse mõistmine või tehisaju loomine oleks nii lihtne, kui me seda tahaks, kuid siiski tundub selle teadmise kasutamine surma pikemaks edasi lükkamiseks suhteliselt lihtsam ja saavutatavam võrreldes teiste mõeldavate võimalustega.
28 November 2009
27 November 2009
Perifeersest närvisüsteemist
Perifeerne närvisüsteem koosneb nendest närvidest ja neuronite kogumikest (ganglionid), mis jäävad väljapoole aju ning seljaaju.
Nägemismeel

Inimesel on 5-6 miljonit koonusrakku ja 120-140 miljonit kepprakku. Lisaks osaleb 800 000- 1 000 000 ganglionrakku nende regulatsioonis enne 1,2 miljoni raku kaudu (silma kohta) signaalide ajusse saatmist. Keppraku aktiveerimiseks piisab ühest footonist ning koonusraku aktiveerimiseks kulub 100 footonit.
Nägemiseks vajalikud kepp- ja koonusrakud reageerivad silma tagaküljel üldvalgusele ja värvidele. Kepprakud võtavad signaale vaateväljast laialdaselt, kuid koonusrakud on koondunud pigem vaatevälja keskossa. Koonused vajavad aktiveerumiseks rohkem valgust ja seetõttu oleme piisava hämaruse korral värvipimedad. Kepikesed algatavad närviimpulsi teket järgmises rakus, kui retinool muudab opsiinis valgusega reageerides kuju ja eraldub opsiini valgust. Opsiin, mis on G-valgu seoseline retseptorvalk, põhjustab aktiveerudes G-valgu vabastamisega järjest mitme biokeemilise protsessi läbiviimist, mis lõppeb kepikese rakumembraanis olevate ioonkanalite sulgemisega, mis vähendab kepprakkudes impulsi teket. See omakorda põhjustab aktsioonipotentsiaali teket järgmises rakus ja alles siis nägemissignaali edasi kandumist. Kui ioonkanalid sulguvad nägemisrakkudes, siis väheneb ka kaltsiumiioonide sissevool ja kaltsiumist sõltuvad valgud rakus taastavad G-valgu signalisatsiooni tagajärjel lagundatud ained avades sellega jälle ioonkanalid. Kui Opsiini valgud asuvad ketaste laadi struktuuride membraanis ja aktiveerumisel lisatakse sellele fosfaatgruppe, mis soodustab retseptori toimet blokeeriva arrestiini kinnitumist opsiinile ja see on üks põhjus, miks silmad harjuvad eredama valgusega. Arvestades nägemise kiirusest on otseselt näha, et need 5 molekulaarbioloogilist protsessi suudavad päris ruttu ära toimuda.
 Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.
Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.
Enne ajule saatmist suudavad esmastele nägemisrakkudele järgnevad rakud naabrite inhibeerimisega mõjutada signaalide edasikandumist. Üldiselt pääsevad natuke tugevamalt edasi piirid või kontrastselt erineva piiriala, mis teeb sama välja. Lisaks lihtsustab see liikumise märkamist. Näidet sellest protsessist näeb selle saidi alaosas.
Neurotransmitterid. Glutamaati leidub ja see paistab muidu kõige olulisemana, arvestades kui tavaline see on sensoorsetes neuronites ja teisteks olulistemateks neurotransmitteriteks silmanärvi neuronitel on inhibeerivad glütsiin ja GABA. Teiseks ei paista, et leidub teisi aktiveerivaid signaali esilekutsujaid silmas, kuigi inhibeerivat kohalikku kontrolli esineb. Sellega seondumiseks on NMDA ja AMPA retseptoreid.
Silmanärvid ise viivad inimesel signaale ajusse rohkem kui miljoni aksoniga.
 Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala).
Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala).
Haistemeelest
Haistenärvid sisaldavad G-valgu seoselisi retseptorvalke, mis aktiveeruvad, kui neile kinnituvad kindlate omadustega molekulid. Tekkinud signaalid kanduvad inimesel aju all olevatesse piklikesse haistesibulatesse (olfactory bulb).
 Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.
Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.
Glutamaadi retseptoritest sisaldab lõhnasibul metabotroopseid glutamaadi, AMPA ja NMDA retseptoreid. GABA jaoks on GABA A ja GABA B retseptorid. Aju poolt on reguleeritud atsetüülkoliini, noradrenaliini, dopamiini ja serotoniini eritavate ühenduste poolt. CB1 retseptorid suudavad mõjutada lõhnasibula neuronite aktiivsust.
Maitsmisest
 Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.
Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.
Keele maitsepungad sisaldavad G-valgu seoselisi aineid struktuuri järgi ära tundvaid retseptoreid. Aktiveerimisel tekitavad signaali kaaliumiioonide väljavoolu takistamise ja positiivse laenguga ioonide sissevoolu algatamisega. Signaalide maitsepungadest edasi levimine käib ähmasemal viisil, sest rotil näiteks osaleb selles 3 kraniaalnärvi paari. Lisaks 9. kraniaalnärvi paarile on keelega ühenduses lisaks vagaalnärv ja näonärv. Nende maitsele reageerivad osad on kõik sama ajutüve tuumaga ühenduses ja sellest edasi jõuavad need muuhulgas ventraal posterioorse mediaalse taalamuse tuuma kaudu insulani. Selles töös mainiti möödaminnes, et loodi transgeenne hiir, kellel oli magusatajus osalev T1R1 retseptor vahetatud opioidi retseptoriga, misjärel reageerisid nad sellega seonduvat ainet maitstes nagu oleks see magus. Tavahiired nii ei reageerinud. Maitseretseptoreid leidub närilistel suuremal hulgal ka suulaes (isiklikult tundsin suulae tagumise pehmema osaga mee maitset selgelt) ja kurgus ning tugevamamaitselisi toite süües jääb meil järelmaitse tuntavaks palju laiemas piirkonnas, kui ainult keelel.
Signaalide edasikandumises osalevad glutamaadi kainaat, AMPA ja NMDA retseptorid. GABA retseptorid on olemas inhibeeriva rolliga.
Kuulmine
Väliskeskkonnast keskkõrva kandunud õhuvõnkumised põhjustavad kuulmekarvade (mehhanosensoritena toimivad rakud) aktiveerumist. Labürindi seinal olevad vestibulaarsed (tasakaalu) ripsrakud reageerivad 3 eri orientatsioonis olevas poolringi kujulises ruumis oleva vedeliku liikumisele keha liikumise tõttu ja aktiveerivad enda küljes oleva neuroni.
Ajusse viivad auditoorseid signaale umbes 30 000 aksonit. Glutamaadi retseptorid olulised kuulmissignaalide vahendajad. Tasakaaluorgani signaalide vahendajaks glutamaadi AMPA, NMDA ja metabotroopsed retseptorid.
Lapse kuulmisvahemik on 20-20 000 Hz ja vanadel inimestel võib see olla 50-8000 Hz. Ning nende 30 000 neuroni vahendatud signaalid jõuavad 570 000 mediaalse genikulaattuuma neuroni vahendusel auditoorsele korteksile, milles on ~100 miljonit neuronit.
Kuulmiskahjustused tekivad pikajalise ~90 db helitugevuse korral ja valu algab 130 db helide korral.
Lihaskontroll
Tahtlik lihaskontroll toimub alfa mootorneuronite abil ja selleks kasutatakse atsetüülkoliini, mis lihasrakkudel olevate nikotiinsete atsetüülkoliini retseptoritega seondub. Nende aktivatsioon põhjustab naatriumi ioonide sissevoolu ja lihasraku mebraanipotentsiaali muutust sarnaselt neuronitega. Raku siselaendu muutus põhjustab kaltsiumi ioonide järsku vabanemist rakusisestest varudest ning kuigipalju ka rakuvälisest keskkonnast sisse tulekut, mis omakorda algatab mootorvalkude poolset energia kulutamist ja lihasraku kokkutõmmet. Atsetüülkoliin lagundatakse kiiresti või kandub passiivselt rakuvälises keskkonnas mujale. Nikotiinsete atsetüülkoliini antagonistid halvavad tahtlikule kontrollile alluvad lihased ja neid kasutatakse kirurgias patsiendi ajutiseks halvamiseks, sest üldnarkoosis olles võib muidu kerget liikumist ette tulla ja teiseks lõdvestab see lihaseid, lihtsustades ligipääsu lihaste taga olevale.
Oksütotsiinist
Oksütotsiin on vajalik piima eritumiseks ja see on osaliselt neuroloogiliselt reguleeritud. Kui nibu stimuleeritakse puudutusega, siis kanduvad signaalid sellest hüpotaalamusse, mis põhjustab oksütotsiini eritumist ajuripatsist verre. Inimeste puhul võib seetõttu mööduda 30-60 sekundit, enne kui piim hakkab imetamisel erituma. Seal artiklis kirjutati, et kuna aju kontrollib oksütotsiini vabanemist, siis võib selle vabanemine alata ka enne imetamist nagu näiteks nutu kuulmisel.
Mehhano- ja termoretseptorid nahas

Puudutustele ja survele reageerivad mehhanoretseptorid põhjustavad aktsioonipotentsiaali, kui kuju muutus selles olevale neuronile muudab ümbritseva vedeliku survet ja mõjutab ioonide suhet vedeliku sissevoolu mõjutamisega. Tavaliselt on valuretseptoriteks ülaloleval pildil asuvad vabad närvilõpmed, kuid ka need reageerivad puudutustele ja temperatuurile mingil määral.
Temperatuuritajus osalevad temperatuuritundlikud ioonkanalid mis võivad reageerida ka farmakoloogilisele aktiveerimisele. Erinevad temperatuuriretseptorid võivad osaleda neutraalses ja valutekitavalt intentsiivses temperatuuritajus. Mentool aktiveerib külmaretseptorid ja kapsaitsiin tsillipiprast aktiveerib kuumaretseptoreid. Viimane tekitab ka valutaju.
Nervus vagus
N. vagus e. uitnärv või vagaalnärv on kraniaalnärv, mis on nii motoorse kui sensoorse toimega. Teised kraniaalnärvid piirduvad tavaliselt pea ja kaelapiirkonnaga, kuid n. vagus saadab ühendusi jämesooleni (mitte lõpuni). Ühe elutähtsama funktsiooniga saadab see ajust südamesse aeglustavaid ühendusi. Südant stimuleerib närvisüsteem veidi kaudsemalt adrenaliiniretseptorite aktiveerimisega adrenaliini vabastamisega mujalt. Uitnärv vajab atsetüülkoliini oma inhibeerivate signaalide saatmiseks ja seetõttu suudavad atsetüülkoliini blokeerivad ained pulsi väga kiireks ajada. Elustamisel kasutatakse südame liikumasaamiseks või lihtsalt kiirendamiseks adrenaliini või atropiini (atsetüülkoliini blokeerija). Mõlemal on potentsiaali liiga kiire pulsi tekitamisel surmav olla.
Pudendaalnärv
Pudendaalnärv on mõlema soo esindajate genitaalide ja lähiümbruse närv, mis omab sensoorseid ja motoorseid ühendusi. Motoorsed reguleerivad põie ja päraku lihaseid. Sensoorsed pudendaalsed ühendused kannavad paaritumises olulisi signaale sh. mõnu selgroonärvi kaudu ajusse. Signaalid kanduvad edasi peamiselt glutamaadi vahendusel. Selgroonärvist tagasi tulevad ühendused, mis põhjustavad kohalikku verevoolu tõusu ning erutusmuudatusi atsetüülkoliini poolse aktivatsiooniga. Noradrenaliin mõjub erutust inhibeerivana neurotransmitterina. Mingi roll on selliste signaalide kandumisel ka vagaalnärvil.
Selgroonärv
 Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.
Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.
 Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.
Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.
Nucleus proprius kannab edasi sensoorset infot puudutuste, jäsemete asukohtade, temperatuuri ja valu kohta kohta. Asub lamina III-V piirkonnas. Dorsaalne tuum (lamina VI-VII piirkonnas) vahendab alateadlikke signaale kehaosade asukohtadest cerebellumile ja vigastus põhjustab ataksiat ning kehaosade kordinatsioonihäired. Lisaks vahendavad VI-VII kihid signaale somatosensoorsele korteksile. Signaalid puudutustest ja asukohtadest liiguvad kiirusega 35-120 m/s.
Lamina VII-IX vahendavad motoorseid signaale lihastesse. VII e. intermediolateraaltuum reguleerib lisaks autonoomsetele lihastele veel sümpateetilist närvisüsteemi. Umbes IX piirkonnas asuv Onuf'i tuum saadab isastel rohkem motoorseid ühendusi genitaalidesse kui emastes.
Lamina X ümritseb selgroovedelikku sisaldavat tsentraalkanalit, ühendab selgroonärvi pooli ja vahendab kehast pärinevaid signaale ning reguleerib näiteks põie lihaseid, kuid ülevaatlikumat infot on raske leida. Osaleb parasümpateetilise närvisüsteemi aktiveerimisel ja sealhulgas tegeleb seksuaalses erutuses genitaalide verevoolu regulatsiooniga (mitte erektsioonis).
Lisaks liiguvad signaalid mõlemas suunas ka valges massis (ainult mitte nii korrapäraselt ja funktsionaalselt üleminevalt) ja seal paistab ühe olulisema ühendusena teadvuse jaoks spinoretikulaarne trakt, mis kannab kehast valusignaale ja lõpetab oma ühendused retikulaarses formatsioonis.
Nägemismeel

Inimesel on 5-6 miljonit koonusrakku ja 120-140 miljonit kepprakku. Lisaks osaleb 800 000- 1 000 000 ganglionrakku nende regulatsioonis enne 1,2 miljoni raku kaudu (silma kohta) signaalide ajusse saatmist. Keppraku aktiveerimiseks piisab ühest footonist ning koonusraku aktiveerimiseks kulub 100 footonit.
Nägemiseks vajalikud kepp- ja koonusrakud reageerivad silma tagaküljel üldvalgusele ja värvidele. Kepprakud võtavad signaale vaateväljast laialdaselt, kuid koonusrakud on koondunud pigem vaatevälja keskossa. Koonused vajavad aktiveerumiseks rohkem valgust ja seetõttu oleme piisava hämaruse korral värvipimedad. Kepikesed algatavad närviimpulsi teket järgmises rakus, kui retinool muudab opsiinis valgusega reageerides kuju ja eraldub opsiini valgust. Opsiin, mis on G-valgu seoseline retseptorvalk, põhjustab aktiveerudes G-valgu vabastamisega järjest mitme biokeemilise protsessi läbiviimist, mis lõppeb kepikese rakumembraanis olevate ioonkanalite sulgemisega, mis vähendab kepprakkudes impulsi teket. See omakorda põhjustab aktsioonipotentsiaali teket järgmises rakus ja alles siis nägemissignaali edasi kandumist. Kui ioonkanalid sulguvad nägemisrakkudes, siis väheneb ka kaltsiumiioonide sissevool ja kaltsiumist sõltuvad valgud rakus taastavad G-valgu signalisatsiooni tagajärjel lagundatud ained avades sellega jälle ioonkanalid. Kui Opsiini valgud asuvad ketaste laadi struktuuride membraanis ja aktiveerumisel lisatakse sellele fosfaatgruppe, mis soodustab retseptori toimet blokeeriva arrestiini kinnitumist opsiinile ja see on üks põhjus, miks silmad harjuvad eredama valgusega. Arvestades nägemise kiirusest on otseselt näha, et need 5 molekulaarbioloogilist protsessi suudavad päris ruttu ära toimuda.
 Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.
Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.Enne ajule saatmist suudavad esmastele nägemisrakkudele järgnevad rakud naabrite inhibeerimisega mõjutada signaalide edasikandumist. Üldiselt pääsevad natuke tugevamalt edasi piirid või kontrastselt erineva piiriala, mis teeb sama välja. Lisaks lihtsustab see liikumise märkamist. Näidet sellest protsessist näeb selle saidi alaosas.
Neurotransmitterid. Glutamaati leidub ja see paistab muidu kõige olulisemana, arvestades kui tavaline see on sensoorsetes neuronites ja teisteks olulistemateks neurotransmitteriteks silmanärvi neuronitel on inhibeerivad glütsiin ja GABA. Teiseks ei paista, et leidub teisi aktiveerivaid signaali esilekutsujaid silmas, kuigi inhibeerivat kohalikku kontrolli esineb. Sellega seondumiseks on NMDA ja AMPA retseptoreid.
Silmanärvid ise viivad inimesel signaale ajusse rohkem kui miljoni aksoniga.
 Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala).
Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala). Haistemeelest
Inimesel on 12 miljonit haistenärvi 10 ruutsentimeetrisel pinnal (koeral võib olla 1-4 miljardit lõhna tajuvat neuronit 170 ruutsentimeetril).
Haistenärvid sisaldavad G-valgu seoselisi retseptorvalke, mis aktiveeruvad, kui neile kinnituvad kindlate omadustega molekulid. Tekkinud signaalid kanduvad inimesel aju all olevatesse piklikesse haistesibulatesse (olfactory bulb).
 Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.
Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.Glutamaadi retseptoritest sisaldab lõhnasibul metabotroopseid glutamaadi, AMPA ja NMDA retseptoreid. GABA jaoks on GABA A ja GABA B retseptorid. Aju poolt on reguleeritud atsetüülkoliini, noradrenaliini, dopamiini ja serotoniini eritavate ühenduste poolt. CB1 retseptorid suudavad mõjutada lõhnasibula neuronite aktiivsust.
Maitsmisest
 Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.
Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.Keele maitsepungad sisaldavad G-valgu seoselisi aineid struktuuri järgi ära tundvaid retseptoreid. Aktiveerimisel tekitavad signaali kaaliumiioonide väljavoolu takistamise ja positiivse laenguga ioonide sissevoolu algatamisega. Signaalide maitsepungadest edasi levimine käib ähmasemal viisil, sest rotil näiteks osaleb selles 3 kraniaalnärvi paari. Lisaks 9. kraniaalnärvi paarile on keelega ühenduses lisaks vagaalnärv ja näonärv. Nende maitsele reageerivad osad on kõik sama ajutüve tuumaga ühenduses ja sellest edasi jõuavad need muuhulgas ventraal posterioorse mediaalse taalamuse tuuma kaudu insulani. Selles töös mainiti möödaminnes, et loodi transgeenne hiir, kellel oli magusatajus osalev T1R1 retseptor vahetatud opioidi retseptoriga, misjärel reageerisid nad sellega seonduvat ainet maitstes nagu oleks see magus. Tavahiired nii ei reageerinud. Maitseretseptoreid leidub närilistel suuremal hulgal ka suulaes (isiklikult tundsin suulae tagumise pehmema osaga mee maitset selgelt) ja kurgus ning tugevamamaitselisi toite süües jääb meil järelmaitse tuntavaks palju laiemas piirkonnas, kui ainult keelel.
Signaalide edasikandumises osalevad glutamaadi kainaat, AMPA ja NMDA retseptorid. GABA retseptorid on olemas inhibeeriva rolliga.
Kuulmine
Väliskeskkonnast keskkõrva kandunud õhuvõnkumised põhjustavad kuulmekarvade (mehhanosensoritena toimivad rakud) aktiveerumist. Labürindi seinal olevad vestibulaarsed (tasakaalu) ripsrakud reageerivad 3 eri orientatsioonis olevas poolringi kujulises ruumis oleva vedeliku liikumisele keha liikumise tõttu ja aktiveerivad enda küljes oleva neuroni.
Ajusse viivad auditoorseid signaale umbes 30 000 aksonit. Glutamaadi retseptorid olulised kuulmissignaalide vahendajad. Tasakaaluorgani signaalide vahendajaks glutamaadi AMPA, NMDA ja metabotroopsed retseptorid.
Lapse kuulmisvahemik on 20-20 000 Hz ja vanadel inimestel võib see olla 50-8000 Hz. Ning nende 30 000 neuroni vahendatud signaalid jõuavad 570 000 mediaalse genikulaattuuma neuroni vahendusel auditoorsele korteksile, milles on ~100 miljonit neuronit.
Kuulmiskahjustused tekivad pikajalise ~90 db helitugevuse korral ja valu algab 130 db helide korral.
Lihaskontroll
Tahtlik lihaskontroll toimub alfa mootorneuronite abil ja selleks kasutatakse atsetüülkoliini, mis lihasrakkudel olevate nikotiinsete atsetüülkoliini retseptoritega seondub. Nende aktivatsioon põhjustab naatriumi ioonide sissevoolu ja lihasraku mebraanipotentsiaali muutust sarnaselt neuronitega. Raku siselaendu muutus põhjustab kaltsiumi ioonide järsku vabanemist rakusisestest varudest ning kuigipalju ka rakuvälisest keskkonnast sisse tulekut, mis omakorda algatab mootorvalkude poolset energia kulutamist ja lihasraku kokkutõmmet. Atsetüülkoliin lagundatakse kiiresti või kandub passiivselt rakuvälises keskkonnas mujale. Nikotiinsete atsetüülkoliini antagonistid halvavad tahtlikule kontrollile alluvad lihased ja neid kasutatakse kirurgias patsiendi ajutiseks halvamiseks, sest üldnarkoosis olles võib muidu kerget liikumist ette tulla ja teiseks lõdvestab see lihaseid, lihtsustades ligipääsu lihaste taga olevale.
Oksütotsiinist
Oksütotsiin on vajalik piima eritumiseks ja see on osaliselt neuroloogiliselt reguleeritud. Kui nibu stimuleeritakse puudutusega, siis kanduvad signaalid sellest hüpotaalamusse, mis põhjustab oksütotsiini eritumist ajuripatsist verre. Inimeste puhul võib seetõttu mööduda 30-60 sekundit, enne kui piim hakkab imetamisel erituma. Seal artiklis kirjutati, et kuna aju kontrollib oksütotsiini vabanemist, siis võib selle vabanemine alata ka enne imetamist nagu näiteks nutu kuulmisel.
Mehhano- ja termoretseptorid nahas

Puudutustele ja survele reageerivad mehhanoretseptorid põhjustavad aktsioonipotentsiaali, kui kuju muutus selles olevale neuronile muudab ümbritseva vedeliku survet ja mõjutab ioonide suhet vedeliku sissevoolu mõjutamisega. Tavaliselt on valuretseptoriteks ülaloleval pildil asuvad vabad närvilõpmed, kuid ka need reageerivad puudutustele ja temperatuurile mingil määral.
Temperatuuritajus osalevad temperatuuritundlikud ioonkanalid mis võivad reageerida ka farmakoloogilisele aktiveerimisele. Erinevad temperatuuriretseptorid võivad osaleda neutraalses ja valutekitavalt intentsiivses temperatuuritajus. Mentool aktiveerib külmaretseptorid ja kapsaitsiin tsillipiprast aktiveerib kuumaretseptoreid. Viimane tekitab ka valutaju.
Nervus vagus
N. vagus e. uitnärv või vagaalnärv on kraniaalnärv, mis on nii motoorse kui sensoorse toimega. Teised kraniaalnärvid piirduvad tavaliselt pea ja kaelapiirkonnaga, kuid n. vagus saadab ühendusi jämesooleni (mitte lõpuni). Ühe elutähtsama funktsiooniga saadab see ajust südamesse aeglustavaid ühendusi. Südant stimuleerib närvisüsteem veidi kaudsemalt adrenaliiniretseptorite aktiveerimisega adrenaliini vabastamisega mujalt. Uitnärv vajab atsetüülkoliini oma inhibeerivate signaalide saatmiseks ja seetõttu suudavad atsetüülkoliini blokeerivad ained pulsi väga kiireks ajada. Elustamisel kasutatakse südame liikumasaamiseks või lihtsalt kiirendamiseks adrenaliini või atropiini (atsetüülkoliini blokeerija). Mõlemal on potentsiaali liiga kiire pulsi tekitamisel surmav olla.
Pudendaalnärv
Pudendaalnärv on mõlema soo esindajate genitaalide ja lähiümbruse närv, mis omab sensoorseid ja motoorseid ühendusi. Motoorsed reguleerivad põie ja päraku lihaseid. Sensoorsed pudendaalsed ühendused kannavad paaritumises olulisi signaale sh. mõnu selgroonärvi kaudu ajusse. Signaalid kanduvad edasi peamiselt glutamaadi vahendusel. Selgroonärvist tagasi tulevad ühendused, mis põhjustavad kohalikku verevoolu tõusu ning erutusmuudatusi atsetüülkoliini poolse aktivatsiooniga. Noradrenaliin mõjub erutust inhibeerivana neurotransmitterina. Mingi roll on selliste signaalide kandumisel ka vagaalnärvil.
Selgroonärv
 Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.
Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.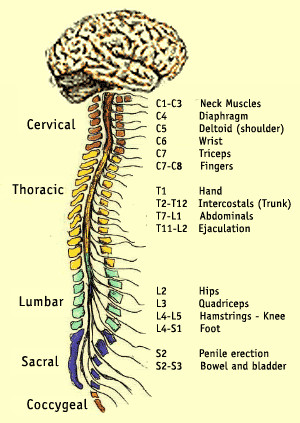{kind=link}
 Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.
Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.Nucleus proprius kannab edasi sensoorset infot puudutuste, jäsemete asukohtade, temperatuuri ja valu kohta kohta. Asub lamina III-V piirkonnas. Dorsaalne tuum (lamina VI-VII piirkonnas) vahendab alateadlikke signaale kehaosade asukohtadest cerebellumile ja vigastus põhjustab ataksiat ning kehaosade kordinatsioonihäired. Lisaks vahendavad VI-VII kihid signaale somatosensoorsele korteksile. Signaalid puudutustest ja asukohtadest liiguvad kiirusega 35-120 m/s.
Lamina VII-IX vahendavad motoorseid signaale lihastesse. VII e. intermediolateraaltuum reguleerib lisaks autonoomsetele lihastele veel sümpateetilist närvisüsteemi. Umbes IX piirkonnas asuv Onuf'i tuum saadab isastel rohkem motoorseid ühendusi genitaalidesse kui emastes.
Lamina X ümritseb selgroovedelikku sisaldavat tsentraalkanalit, ühendab selgroonärvi pooli ja vahendab kehast pärinevaid signaale ning reguleerib näiteks põie lihaseid, kuid ülevaatlikumat infot on raske leida. Osaleb parasümpateetilise närvisüsteemi aktiveerimisel ja sealhulgas tegeleb seksuaalses erutuses genitaalide verevoolu regulatsiooniga (mitte erektsioonis).
Lisaks liiguvad signaalid mõlemas suunas ka valges massis (ainult mitte nii korrapäraselt ja funktsionaalselt üleminevalt) ja seal paistab ühe olulisema ühendusena teadvuse jaoks spinoretikulaarne trakt, mis kannab kehast valusignaale ja lõpetab oma ühendused retikulaarses formatsioonis.
13 November 2009
Cerebellum
 Cerebellum e. väikeaju koosneb kahest väikesest poolkerast ajutüve küljes, mille hinnanguline rakkude arv on suurusele vaatamata tavaliselt suurem, kui ülejäänud ajus kokku. See tegeleb tasakaalumeele andmisega, liigutuste planeerimisega, kõnnakuga ja liigutuste sujuvama/detailsema läbiviimisega olles ajukoore motoorsete alade täiendajaks, kuid paistab omavat rolli ka mõtlemisprotsessides. Cerebellumi enda korteks on sarnase kolmekihilise ehitusega iga funktsiooniga ala puhul ja poolkerasse tulnud info pärineb samast kehapoolest või vastaspoole ajupoolkeralt. Elektriline stimuleerimine ei tekita teadaolevalt liigutusi ega tajusid.
Cerebellum e. väikeaju koosneb kahest väikesest poolkerast ajutüve küljes, mille hinnanguline rakkude arv on suurusele vaatamata tavaliselt suurem, kui ülejäänud ajus kokku. See tegeleb tasakaalumeele andmisega, liigutuste planeerimisega, kõnnakuga ja liigutuste sujuvama/detailsema läbiviimisega olles ajukoore motoorsete alade täiendajaks, kuid paistab omavat rolli ka mõtlemisprotsessides. Cerebellumi enda korteks on sarnase kolmekihilise ehitusega iga funktsiooniga ala puhul ja poolkerasse tulnud info pärineb samast kehapoolest või vastaspoole ajupoolkeralt. Elektriline stimuleerimine ei tekita teadaolevalt liigutusi ega tajusid.  Kahjustused ja häired funktsioonis põhjustavad tasakaaluhäireid, ataksiat (illustratsioon), lihaste lõtvust, raskusi järjestike kiirelt vahelduvate liigutuste läbiviimisel, vähenenud liigutuste täpsust (näiteks sõrmega ninaotsa puudutamisel) ja mittesujuvat silmade liikumist e. nüstagmust (klipp nüstagmusest). Parkinsoni tõve puhul on aktiivsem, kui tervetel kuid basaalganglioni aktiivsus on neil madalam dopamiini puudusest.
Kahjustused ja häired funktsioonis põhjustavad tasakaaluhäireid, ataksiat (illustratsioon), lihaste lõtvust, raskusi järjestike kiirelt vahelduvate liigutuste läbiviimisel, vähenenud liigutuste täpsust (näiteks sõrmega ninaotsa puudutamisel) ja mittesujuvat silmade liikumist e. nüstagmust (klipp nüstagmusest). Parkinsoni tõve puhul on aktiivsem, kui tervetel kuid basaalganglioni aktiivsus on neil madalam dopamiini puudusest.{kind=link}
Ajukoorelt (lõigu allikas) saab signaale peamiselt motoorsetelt aladelt olles mõjutatud veel parietaalsagara ja temporaalsagara poolt. Saadab signaale taalamuse kaudu motoorsetele aladele ja ka prefrontaalsele koorele. Artiklis mainiti, et ponsi tuum, mis vahendab signaale cerebellumiga, on ühendatud mitmekülgsete sensoorsete aladega sh. singulaatkoorega, visuaalse korteksiga ja posterioorse parietaalse loobuliga. Kahepoolse vigastusega indiviidide puhul paistab, et cerebellum on vajalik liigutuste automaatsele tasemele õppimiseks. Kahjustused võivad lisaks põhjustada raskusi ajavahemike hindamisel, reeglipäraste tegevuste õppimisel ja tähelepanu ühelt sensoorselt objektilt teisele viimisel. Autorid mainisid, et selliseid protsesse on seostatud ka prefrontaalse koorega. Ühes uuringus oli 15'l cerebellumi degeneratsiooniga patsientidel suuri raskusi faktiteadmiste omandamisel ja õpitu kasutamisel. Lugemisel ja rääkimisel aktiveerub laialdaselt. Cerebellumi superioorne osa on ühenduses pigem frontaalsagaraga ja inferioorne osa parietaalsagaraga. Pakuti, et ühendused frontaalsagaraga tegelevad hääldusega ja parietaalsagara suunaline liiklus kõnes/lugemises lühimäluga. Parietaalsagaras paistavad asuvana ka kujutlused ja lühiajalised mälestused teistes töödes. Sarnaste sõnade leidmises omasid tähtsat rolli cerebellumi ja temporaalsagara vahelised kahepoolsed ühendused.
 Enamus cerebellumist on kahesuunalise liiklusega ühenduses ajukoorega, võttes vastu mitmekülgset sensoorset infot silmadest, keskkõrvast, nahalt ja liigesepiirkondadest kehaosade suhtelistest asukohtadest. Neocerebellumi (cerebrocerebellum ühe eelneva pildi peal) osa on ühenduses ainult ajukoorde minevate tuumadega ning omab pigem kognitiivset väärtust inimestele. Selgroonärvi kaudu tulnud info kehaosade suhtelistest asukohtadest ja puudutuste saadetakse cerebellumi superioorsel ja inferioorsel poolel asuvatele spetsiifilistele omamoodi "homunkuluse" aladele. Keskkõrvast pärinev vestibulaarne info pea suhtelise asendi kohta gravitatsiooni suhtes suundub peamiselt väikeajusse ja sealgi suhteliselt väiksesse archicerebellumisse.
Enamus cerebellumist on kahesuunalise liiklusega ühenduses ajukoorega, võttes vastu mitmekülgset sensoorset infot silmadest, keskkõrvast, nahalt ja liigesepiirkondadest kehaosade suhtelistest asukohtadest. Neocerebellumi (cerebrocerebellum ühe eelneva pildi peal) osa on ühenduses ainult ajukoorde minevate tuumadega ning omab pigem kognitiivset väärtust inimestele. Selgroonärvi kaudu tulnud info kehaosade suhtelistest asukohtadest ja puudutuste saadetakse cerebellumi superioorsel ja inferioorsel poolel asuvatele spetsiifilistele omamoodi "homunkuluse" aladele. Keskkõrvast pärinev vestibulaarne info pea suhtelise asendi kohta gravitatsiooni suhtes suundub peamiselt väikeajusse ja sealgi suhteliselt väiksesse archicerebellumisse.Cerebellumil paistab olevat mõju emotsioonide üle nagu selle kahjustusega patsientidel leiti. Positiivsed elamused olid neil nõrgemad, ebameeldivamad tugevamad ning aktivatsioon näiteks insulas, prefrontaalses- ja singulaatkoores erines kontrollgrupi omast, kuid siin ei saa välistada puuete mõju üldisele heaolule. Selle töö tugevamaks küljeks oli ülevaate andmine cerebellumi seostest emotsioonidega ja ühendustest emotsioone vahendavate piirkondadega. Ühendusi on hippokampuse, amügdala ja septaaltuumadega. Vigastused posterioorses sagaras või vermises tuimestavad emotsioone. Emotsioone tekitavad elamused toovad fMRI uuringutes kaasa tõusu väikeaju aktiivsuses ja näiteks tugevam reaktsioon naljale tõi selles kaasa suuremat aktiivsuse tõusu.
Saab noradrenaliini locus coeruleus'ilt ja serotoniini dorsaalsest raphe tuumast. Sisaldab noradrenaliini a1 ja a2 retseptoreid. Kannabinoidi CB1 ja adenosiini retseptorid on olemas. GABA ja glutamaat on olulised reguleerijad ning liikumishäireid saab selgemini esile kutsuda glutamaadi blokeerimisega, kui GABA agonistidega. NMDA agonistidega saab deaktiveerida muskariinseid atsetüülkoliini retseptoreid. NMDA antagonistid ise suudavad edukalt esile kutsuda purjusolekule sarnaseid tasakaaluhäireid ja sarnaselt etanoolile põhjustavad need nüstagmust. Samuti on NMDA antagonistidele omane iivelduse teke kui liigutatakse.
Ehitusest. Cerebellumi korteks koosneb kolmest kihist, mis väljapoolt sissepoole lugedes on molekulaarkiht, purkinje rakkude kiht ja graanulrakkude kiht. Aktiveerivad signaalid jõuavad suuremal määral ajust pärinevaid signaale toovate glutamaatsete mossy fiber'ite kaudu graanulrakkudeni, mille hulk ületab ülejäänud neuronite hulga. Graanulrakud saadavad signaali aksoniga edasi molekulaarkihti, kus see hargneb T kujuliselt kaheks ja seal asuvaid selliseid aksoneid nimetatakse paralleelkiududeks. Graanulrakkude osade vahele jäävad purkinje rakud, mis on suuremat sorti neuronid, millele on omaseks ülisuur ja harunenud dendriidivõrastik. Seetõttu on purkinje rakud võimelised signaale vastu võtma rohkem kui 100 000lt rakult. Need harud asuvad umbes tasapinnaliselt paralleelkiudude all ja saavad neilt impulssi tekitavaid glutamaatseid signaale. Purkinje rakud ise sisaldavad samas GABA ja kui impulls tekib, siis saadavad nad seda cerebellumi tuumadele või sellega ühenduses olevatele tuumadele ning need GABA signaalid on teadaolevalt ainsaks väljundiks seal koores toimuvale tegevusele. Vähemal määral on molekulaarkihis Golgi rakke, mis inhibeerivad paralleelkiududelt signaale saades GABA kasutades graanulrakke. Golgi rakud võivad lõpetada inhibeerivate signaalide saatmise kuni sadadeks millisekunditeks saades seda esile kutsuvaid somatosensoorseid signaale sensoorsetelt närvidelt nagu trigerminaalselt närvilt, mis toob sensoorset infot pea piirkonnast, kuid ka jäsemete sensoorsed närvid. Pikema inhibitsiooni tekitajad olid üldiselt suuremad kogemused, mitte pisikese ala sensoorsed elamused. Autorid lisasid, et Golgi rakud töötavad ajust tulnud mossy fiber'ite summutajad, mis lülituvad välja, kui kehast hakkavad signaalid tulema, võimaldades graanulrakkudel aktiveeruda lühikeseks ajaks.
Stellate ja basket rakud inhibeerivad GABA kasutades purkinje rakke. Ajutüve oliivist pärinevad ronivad kiud võivad kasvada ümber purkinje rakkude ja viimaseid stimuleerides suudavad esile kutsuda kiiremat GABA vabanemist. Üldjoonena paistab, et kuhu iganes signaalid jõuavad cerebellumil on vastuseks kohalik GABA signaalide väljasaatmine. Nende GABA signaalide puudus vigastuste tagajärjel ei põhjusta ilmset hüperaktiivsust ega psühholoogilist pingesolekut. Võimalikke tagajärgi arvestades paistab, et see filtreerib midagi üleliigset välja. Inhibeerivate signaalide alateadliku rolli kohta on suhteliselt lihtne oletusi teha, kui tulemuseks peaks suutma liigutusi täpselt läbi teha, oma keha asetust piiritleda või õppida.
Cerebellumist saadavad signaale välja sügavad cerebellaarsed tuumad (DCN). Nendes olevad neuronid aktiveeruvad somatosensoorsest infost sõltuvalt, kuid erinevalt cerebellumi koorest ilma eriti täpse "kaardita", sest enamus DCN neuronitest võivad rotil aktiveeruda nii näo kui esikäppade puudutusel. Autorid kahtlustasid, et need neuronid on vajalikud tegevuste ajastamisel. DCN somatosensoorne kaart on katkendlik, sest naaberalad ei aktiveeru nahal lähedaste alade aktiveerumisel ja sama naha piirkond võib aktiveerida eri DCN alasid. DCN sisaldab vähemalt kolme erinevat neuronipopulatsiooni. Suured glutamaatsed neuronid ühendavad tuumad mitmete aju osadega nagu taalamuse ja punase tuumaga. Keskmised GABA saatvad neuronid saadavad oma sisu inferioorsele oliivile. Kolmas rakugrupp sisaldab väikseid neuroneid, mis saadavad inhibeerivaid glütsiini signaale kohalikele rakkudele. Sensoorne info jõuab cerebellumi koorele tõenäoliselt lateraalse retikulaarse formatsiooni ja inferioorse tuuma kaudu, mis põhjustavad cerebellumi koore aktiveerumise järel DCN inhibitsiooni.
Kokkuvõtteks on cerebellum pisike võimekas lisand teadvuse jaoks, mis suudab meie keha kiiresti ümbritsevast keskkonnast kogutud infoga tasakaalus hoida. Samuti laseb see meie teadvusel keha kontrollida liigutustele tähelepanu pööramata tehes sellega meie elu palju lihtsamaks ning ühendusi ja vigastusi arvestades suudab see tõenäoliselt ka teisi psüühilisi protsesse cerebellumi jaoks analoogse infotöötlusega kergemaks teha.
06 November 2009
Ajutüvi

Ajutüve roll teadvuse jaoks seisneb peamiselt aju-keha vaheliste signaalide vahendamises ja üldaktiivsuse tugevas regulatsioonis. Koosneb kahest suuremast piirkonnast: ~3 sentimeetrisest medulla oblongata'st ja ~2,5 sentimeetrisest ponsist (eesti keeles ka ajusild), mis ülejäänud aju kaasamata suudavad meid elus hoida südame reguleerimise (kuid süda suudab omal jõul töödata vajades peamiselt adrenaliinseid hormoone. Hingamise alal hoidmises on ajutüvel elutähtis roll. Kontrollivad ka aevastus-, kõha- ja oksereflekse. Lisaks loetakse keskaju tihti ajutüve alla. Viimane on aju suurimaks dopamiiniallikaks (substantia nigra ja ventraalne tegmentaalne ala) ning ühendab ajutüve ülejäänud ajuga. Medullas asuv nn. püramiid on lihastele signaale saatvate neuronite ristumiskoht, misjärel enamus neist läheb edasi ajupoolega vastupidist kehapoolt kontrollima (selliseid poolevahetus kohti on eri meeltel mujalgi). Õhupuudustunde algatajaid on ka medullas ja keskajus. Süsihappegaasist tekkinud süsihape langetab pH'd ja see viib lämbumistunde tekkimisele.
Mittesurmavad vigastused ajutüves on ebatavalised ja võivad põhjustada kaelast allapoole halvatust või locked in sündroomi, kus ei saa vahel isegi silmi liigutada, kuigi oldakse teadvusel. Ponsi ja keskaju piirkonna kahjustused võivad põhjustada koomat ning kuigi kooma võib tekkida teiste piirkondade kahjustustega, on sealsete kahjustuste tagajärjel paranemislootus eriti madal.
Ühel ponsi infarkti saanud mehel tekkis kontrollimatu, kuid emotsioonitu naer. Suurema osa ajast paistis ta sümptomiteta, aga tuli ette vahelduvaid naeru ja nutuhoogusid. Järgmisel päeval alles tekkis lihasnõrkus ja seisund halvenes kuni suri. Kaks patsienti olid peale ponsi ja keskaju infarkti hakanud palju haigutama. Ühel olid haigutushood kontrollimatud ja mõlemad said liigsetest haigutushoogudes aja jooksul lahti.
 Ponsi kuklapoolses küljes asub sinakas locus coeruleus, mis saadab noradrenaliini ajule. Locus coeruleus'i aktiivsus sõltub ärkvelolekust ja aktiivsuse tõus vähendab une hulka. Üheks ööpäevaseks reguleerijaks sellele on hüpotaalamuse suprakiasmaatiline tuum, millest algavad ühendused läbivad ka teisi hüpotaalamuse piirkondi. Veel saab ühendusi amügdalast. Inhibeerib parasümpateetilist närvisüsteemi ajutüve kaudu sh. süljenäärmeid ja pupilli reguleerimist valgusele. Glutamaadi süstimisel LC'sse kiirenes pulss ja tõusis vererõhk. Aktiveerib dorsaalset raphe tuuma. LC ja teiste piirkondade osalust ärkveloleku reguleerimises koos kasutatavate neurotransmitteritega on natuke kujutatud sellel diagrammil.
Ponsi kuklapoolses küljes asub sinakas locus coeruleus, mis saadab noradrenaliini ajule. Locus coeruleus'i aktiivsus sõltub ärkvelolekust ja aktiivsuse tõus vähendab une hulka. Üheks ööpäevaseks reguleerijaks sellele on hüpotaalamuse suprakiasmaatiline tuum, millest algavad ühendused läbivad ka teisi hüpotaalamuse piirkondi. Veel saab ühendusi amügdalast. Inhibeerib parasümpateetilist närvisüsteemi ajutüve kaudu sh. süljenäärmeid ja pupilli reguleerimist valgusele. Glutamaadi süstimisel LC'sse kiirenes pulss ja tõusis vererõhk. Aktiveerib dorsaalset raphe tuuma. LC ja teiste piirkondade osalust ärkveloleku reguleerimises koos kasutatavate neurotransmitteritega on natuke kujutatud sellel diagrammil.Aktiveerimisel muudab ajukoore tegevuse (siin lõigus allikaks) EEG järgi ebasünkroonsemaks ja tekitab hippokampuses teeta rütmi. Inaktivatsioon toob kaasa unele omasemat sünkroonset suure amplituudiga madalasageduslikku tegevust. Sisaldab glutamaati ja enkefaliine. Palju stressis olnud rottidel on maksimumaktiivsus väiksem, kuid samas on nende locus coeruleus tundlikum kortikotropiini vabastamise faktori (CRF) stimulatsiooni poolt, mida hüpotaalamus eritab stressihormoonide vabastamiseks ja toimib LC'le olulise aktiveerijana. Sisaldab tihedalt mu retseptoreid ja on nende agonistide poolt inhibeeritud. CRF toob kaasa pigem ümbrust uurivat erksamat käitumist ja mu retseptori agonistid jätavad tähelepanu üksikumatele tegevustele, kuid need erinevused ei ole väga selged. Mu opioidi agonistide pikaajaline tarbimine võib kasutamise lõppedes suurendada locus coeruleus'i aktiivsust.
Sinna minevates aksonites leidub kappa retseptoreid ja selle agonist dünorfiin põhjustab nendes aktiivsuse vähenemist. Locus coeruleusi sees on eri opioidi retseptorid (mu, kappa, delta) kergelt spetsiifilise asukohamustriga.
Vigastades kassil subcoeruleus'i ala kadus neil REM unele omane lihaslõtvus ja nad hakkasid unenägusid välja elama tõstes pead, liikudes ringi ja üritades näiliselt midagi kinni püüda. Ühes uuringus inimestega, kus magades vahel vigastati liigse aktiivsuse tõttu voodikaaslast, leidus ka neil umbes selle piirkonna kahjustusi, kuid nende kahjustused ulatusid ka mujal. GABA agonistidena toimivate bensodiasepiinidega sai seda probleemi ajutiselt ära hoida. Subcoeruleus'i piirkonnast läksid aksonid muuhulgas retikulaarsesse formatsiooni. Hippokampuse teeta laineid esile kutsuda precoeruleus'i stimuleerimisega glutamaadi retseptoritega.
Periakveduktaalne hall mass (pildil hall piirkond selgroovedelikku sisaldava "akvedukti" ümber) on valusignaale vahendav ja reguleeriv piirkond keskajus.
 Saab signaale valusignaale toovatelt aksonitelt, prefrontaalselt korteksilt, hüpotaalamuselt ja amügdalalt. Tõenäoline opioidide valuvaigistava toime läbiviija. Sisaldab suures koguses endorfiine ja enkefaliine koos vajalike retseptoritega. Naerugaasi toimet seal sai blokeerida nii opioidi antagonisti kui NO inhibiitori andmisel. Inimestele annab selle piirkonna elektriline stimuleerimine tugevat leevendust kroonilisele valule. Stimulatsioonijärgne valuvaigistus kestab kuni 5 minutit. Liiga tihedal stimuleerimisel kaob valu vaigistav toime ja elektrilist valuvaigistust saab blokeerida opioidi antagonistidega. Lisaks vahendab see, koos mõne teise keskaju alaga, REM und, mille osakaal suureneb tugevalt, kui anda GABA agonisti seal tihedalt esinevatele GABA retseptoritele, mida teistel keskaju REM une algatajates nii palju ei olnud.
Saab signaale valusignaale toovatelt aksonitelt, prefrontaalselt korteksilt, hüpotaalamuselt ja amügdalalt. Tõenäoline opioidide valuvaigistava toime läbiviija. Sisaldab suures koguses endorfiine ja enkefaliine koos vajalike retseptoritega. Naerugaasi toimet seal sai blokeerida nii opioidi antagonisti kui NO inhibiitori andmisel. Inimestele annab selle piirkonna elektriline stimuleerimine tugevat leevendust kroonilisele valule. Stimulatsioonijärgne valuvaigistus kestab kuni 5 minutit. Liiga tihedal stimuleerimisel kaob valu vaigistav toime ja elektrilist valuvaigistust saab blokeerida opioidi antagonistidega. Lisaks vahendab see, koos mõne teise keskaju alaga, REM und, mille osakaal suureneb tugevalt, kui anda GABA agonisti seal tihedalt esinevatele GABA retseptoritele, mida teistel keskaju REM une algatajates nii palju ei olnud.Stressis olles tõuseb seal endokannabinoidide tase ja CB1 agonistide lisamisel sinna saab nõrgestada valu. Emastel rottidel liigub sealt selgroo suunas rohkem ühendusi ja morfiini toimel aktiveerus neil isastega võrreldes vähem selliseid neuroneid. Need aktiveerivad selgroonärvis valusignaale vahendavates alades olevaid inhibeerivaid neurotransmittereid eritavad neuronid. Nende neuronite aktiveerumise kaudu paistab, et tekibki periakveduktaalse ala valuvaigistav toime. Mu retseptorite hulk oli isastel kõrgem. Ka inimeste puhul paistab nii, et naised saavad opioididest vähem abi valu vastu. Suurimat erinevust leidnud uuringu järgi pidi naistele andma sama efekti saavutamiseks 30% rohkem morfiini kui meestele. Progestorooni tootmise langus hilise diestruse faasis toob kaasa GABA A retseptorite tootmise tõusu, millest tingitud aktiivsuse vähenemine võib valutaju tugevdada. 5-HT2C antagonistide doseerimisel sinna vähenes kassil kaitsev agressiivsus.
Suurimaks struktuuriks ajutüves on retikulaarne formatsioon, mis läbib seda medullast keskajuni. See on teadvuse seisukohalt ülioluline une-ärkveloleku reguleerija/alalhoidja ning tihti mainitakse selle juures retikulaarset aktivatsiooni süsteemi, mille all mõeldakse selle võimet aju teadvusele tuua, kuid ka magama panna. Retikulaarne formatsioon saab sensoorseid signaale seljaajult ja eri meeltelt ning annab neid muuhulgas edasi taalamusele. Retikulaarse aktivatsiooni tuumade kahjustusi keskaju ja ponsi retikulaarses formatsioonis seostatakse kooma ja vegetatiivse seisundiga. Selle piirkonna vigastusest tingitud kooma lõppeb kiiremini ja palju kindlamalt surmaga, kui teistest kahjustustest tekkinud kooma. Samuti võis vegetatiivne seisund kaasneda, kui patsiendil oli vigastus taalamuses või taalamuse ja retikulaarse formatsiooni vahelises ühenduses.
On üks (kahest, kui see artikkel midagi välja ei jätnud) PGO ajulaine tekitajast, mis pärinevad ponsist, läbivad lateraalse genikulaat tuuma ja jõuavad kuklasagarale REM une ajal. Lateraal genikulaatse tuuma läbimisel kattusid need lained REM une ajal toimuvate silmaliigutustega (selle piirkonna läbimine ise ei tohiks tekitada silmaliigutusi, kuid juhuslikult mõõdeti need koos toimuvana ja PGO lained läbivad muidki piirkondi). Need lained paistavad une ajal peamised signaalisaatjad kuklasagarale une ajal (REM uni on tavaline unenägude nägemise aeg). Unespindlite teke une ajal seondus selles artiklis viidatud uuringu järgi retikulaarses formatsioonis samaaegselt tekkinud takistustega kehalt tulnud signaalide vahendusel.
 Ponsi jääva retikulaarse (tõlkes võrkja) formatsiooni oraalne osa tekitab GABA taseme tõstmisel ärkvelolekut. Hüpotaalamusest tulev hüpokretiin-1 põhjustas (lõigu põhiallikas) seal rakkudevahelises keskkonnas GABA taseme tõusu ja ärkvelolekut. Samas töös GABA sünteesi inhibiitori lisamisel vähenes ärkveloleku aeg ning pikenesid REM ja NREM une faasid. Autorid lisasid, et seetõttu ei ole GABA agonistid head unetusevastased ravimid. Hüpokretiini enda puudulikkust sh. mutatsioone selle retseptoris seostati narkolepsiaga inimestel ja koertel. Hüpokretiini signalisatsiooni rikkumisel mutantsetel hiirtel saadi isendid, kellel olid narkolepsia sümptomid. Narkolepsia vastane ravim modanifil aktiveeris hüpokretiini neuroneid.
Ponsi jääva retikulaarse (tõlkes võrkja) formatsiooni oraalne osa tekitab GABA taseme tõstmisel ärkvelolekut. Hüpotaalamusest tulev hüpokretiin-1 põhjustas (lõigu põhiallikas) seal rakkudevahelises keskkonnas GABA taseme tõusu ja ärkvelolekut. Samas töös GABA sünteesi inhibiitori lisamisel vähenes ärkveloleku aeg ning pikenesid REM ja NREM une faasid. Autorid lisasid, et seetõttu ei ole GABA agonistid head unetusevastased ravimid. Hüpokretiini enda puudulikkust sh. mutatsioone selle retseptoris seostati narkolepsiaga inimestel ja koertel. Hüpokretiini signalisatsiooni rikkumisel mutantsetel hiirtel saadi isendid, kellel olid narkolepsia sümptomid. Narkolepsia vastane ravim modanifil aktiveeris hüpokretiini neuroneid.Nii inimestel kui loomadel põhjustas seal atsetüülkoliini tõstmine NREM une ajal REM une faasi minekut, kuid sama doosi andmisel REM unne üleminekul põhjustas ärkamist. Kassidel ärkveloleku ajal atsetüülkoliini tõstmisel tekkis dissotseerunud seisund, mis meenutas kerge lihaste lõtvuse poolest REM und, kuid nad olid ärkvel ja jälitasid silmadega liikumist. Sarnaselt põhjustas GABA tõus ärkvelolekut ka keskaju retikulaarses formatsioonis ja hüpotaalamuse preoptilisel/anterioorsel alal. Hüpotaalamuse posterioorses osas põhjustas GABA und nagu tavadoseerimisel. Mujal ajus toimib hüpokretiin stimuleerivalt noradrenaliini, serotoniini ja atsetüülkoliini vabastamisel. GABA A antagonisti andmisel tõusis atsetüülkoliini hulk. Adenosiini A2A agonisti andmisel vabanes rohkem atsetüülkoliini ja kutsuti esile REM und.
Üldnarkoosis isofluraaniga vähenes GABA tase retikulaarse formatsiooni oraalses alas. Autorid pakkusid seda toimemehhansimiks, kuid eelmise lõigu juttu toetavaid artikleid oli umbes kümme korda rohkem ja tõin pigem näiteks sellest, et teadustööde puhul on tähtis võimalikult paljude tööde najal järeldusi teha. Ka morfiini puhul leiti GABA vähenemist seal ning autorid pidasid seda põhjuseks sellele, miks morfiin tekitab sügavama une vähesust.
Ühes katses lisati elektroodid medullas asuvasse retikulaarsesse formatsiooni ja selle stimuleerimisel üldnarkoosi ajal 50-300 Hz juures vähenes ajukoore EEG'l suureamplituudiliste ajulainete hulk ja tugevus. 200-300 hertsiline simulatsioon eriti vähendasid delta (0,5-4 Hz) ja teeta sagedusel (4-8 Hz) lainete hulka. Tekkisid mõõdetavad umbes gamma sagedusel ajulained 22-50 hertsi juures. Sellele ajupiirkonnale olid omaseks hästi suured neuronid. Pildil on näha stimulatsiooni mõju laineamplituudile ja off-target oli kontrollgrupp, kellel stimuleeriti veidi kõrvalist piirkonda, et näiteks ei saaks EEG tulemust puhtalt elektrilise stimulatsiooni taustmüraks pidada.
Retikulaarse formatsiooni keskaju sisene osa sisaldab aju peamist serotoniiniallikat, mida nimetatakse dorsaalseks raphe tuumaks. Inimestelt leiti dorsaalsest raphe tuumast lahkamise järgselt ka neuropeptiid P'd, kuid serotoniini sisaldavate neuronite hulk oli 2 korda suurem. Amügdalasse saadavad DRT'st aksoneid 10% sealsetest serotoniini sisaldavatest neuronitest.
Kokkuvõtlikult ning lihtsustatult on sügava une tekitajad illustreeritud siin ja ärkvelolekus osalev aktiveerumise ahel siin.
Kraniaalnärvid vahendavad mitmete meelte (põhiliselt pea piirkonna meelte) infot ja kuna nad ei sõltu otseselt seljaajust, ei teki seljaaju või selgroonärvi vigastuste puhul vähemalt nende vahendatud meelte ja lihaste kontrolli kadumist. 12st kraniaalnärvi paarist 10 on ühendatud ajutüvega. Näiteks okserefleksi korral saab IX närvipaar sensoorse närvina signaale kurgust, kõrist ning keele ja suulae tagaosast. Signaalid jõuavad selgroonärvi trigeminaalse tumani (Spinal trigeminal nucleus või lihtsalt Sp5). See mõjutab nucleus ambiguus'i, mis algatab vagaalnärvi motoorsete ühenduste kaudu okserefleksi lihastõmblused.

4. juuni 2010 täiendus

 Silmaliigutusi reguleerivad 3 kraniaalnärvi paari. Number näitab milline närv liigutab silma selles suunas ja tume silmanurk tähistab ninapoolset külge.
Silmaliigutusi reguleerivad 3 kraniaalnärvi paari. Number näitab milline närv liigutab silma selles suunas ja tume silmanurk tähistab ninapoolset külge.Neuronite hulga võtsin siit.
Okulomotoorsel (3.) kraniaalnärvil on 25 000-35 000 neuronit silma liigutuseks osades suunades.
4. kraniaalnärv kontrollib ühte silmalihast 2000-3500 neuroniga ning sama palju on selle tuumas.
5. kraniaalnärv e. trigeminaalne närv saab näost ja suust sensoorset infot puudutuse ja valu kohta 140 000 neuroniga. Selle motoorsed ühendused mälumiseks ja neelamiseks sisaldavad 8100 rakku.
6. kraniaalnärv liigutab silma ühe lihase kontrolliga ninast eemale 3700 rakuga.
7. kraniaalnärv e. näonärv kontrollib näoilmeid, pisaraeritust, süljeeritust, parasümpateetiliselt ninaõõnsust ja vahendab osaliselt maitsesignaale 9000-10 000 neuroniga. Näonärvi tuumas on 7000 rakku ja selle pikkus on 2-5,6 mm. Paistab, et näoilme kontrolliks on kordades vähem neuroneid kasutusel, kui silmade liigutamiseks.
8. paar vahendab keskkõrvast pärinevaid pea orientatsiooni signaale.
9. kraniaalnärv saab signaale keele tagumiselt kolmandikult ja vahendab öökimisrefleksi algatavaid sensoorseid signaale.
10. ehk vagaalnärv suudab tekitada oksendamist ning vahendab siseorganite sensoorseid signaale.
11. kraniaalnärv kontrollib kahte kaelalihast ning on suuremaltjaolt ühinenud vagaalnärviga (ajutüve lähedal liitub) vahendades vagaalnärviga samadest piirkondadest tulnud signaale.
12. kraniaalnärvi tuumas on 4500-7500 neuronit ning see vahendab suu tagaosast pärinevaid maitse- ja muid sensoorseid signaale ning on peamine keelelihaste kontrollija.