Nägemismeel

Inimesel on 5-6 miljonit koonusrakku ja 120-140 miljonit kepprakku. Lisaks osaleb 800 000- 1 000 000 ganglionrakku nende regulatsioonis enne 1,2 miljoni raku kaudu (silma kohta) signaalide ajusse saatmist. Keppraku aktiveerimiseks piisab ühest footonist ning koonusraku aktiveerimiseks kulub 100 footonit.
Nägemiseks vajalikud kepp- ja koonusrakud reageerivad silma tagaküljel üldvalgusele ja värvidele. Kepprakud võtavad signaale vaateväljast laialdaselt, kuid koonusrakud on koondunud pigem vaatevälja keskossa. Koonused vajavad aktiveerumiseks rohkem valgust ja seetõttu oleme piisava hämaruse korral värvipimedad. Kepikesed algatavad närviimpulsi teket järgmises rakus, kui retinool muudab opsiinis valgusega reageerides kuju ja eraldub opsiini valgust. Opsiin, mis on G-valgu seoseline retseptorvalk, põhjustab aktiveerudes G-valgu vabastamisega järjest mitme biokeemilise protsessi läbiviimist, mis lõppeb kepikese rakumembraanis olevate ioonkanalite sulgemisega, mis vähendab kepprakkudes impulsi teket. See omakorda põhjustab aktsioonipotentsiaali teket järgmises rakus ja alles siis nägemissignaali edasi kandumist. Kui ioonkanalid sulguvad nägemisrakkudes, siis väheneb ka kaltsiumiioonide sissevool ja kaltsiumist sõltuvad valgud rakus taastavad G-valgu signalisatsiooni tagajärjel lagundatud ained avades sellega jälle ioonkanalid. Kui Opsiini valgud asuvad ketaste laadi struktuuride membraanis ja aktiveerumisel lisatakse sellele fosfaatgruppe, mis soodustab retseptori toimet blokeeriva arrestiini kinnitumist opsiinile ja see on üks põhjus, miks silmad harjuvad eredama valgusega. Arvestades nägemise kiirusest on otseselt näha, et need 5 molekulaarbioloogilist protsessi suudavad päris ruttu ära toimuda.
 Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.
Koonusrakkudes on valgusretseptorid sarnaselt ketasjates moodustistes, aga kergete erinevuste tõttu aminohappelise järjestuse reageerivad need kergemini spetsiifilistele lainepikkustele, muutes retinooli kuju eelistatult teatud värvusega lainepikkuseid vastu võttes.Enne ajule saatmist suudavad esmastele nägemisrakkudele järgnevad rakud naabrite inhibeerimisega mõjutada signaalide edasikandumist. Üldiselt pääsevad natuke tugevamalt edasi piirid või kontrastselt erineva piiriala, mis teeb sama välja. Lisaks lihtsustab see liikumise märkamist. Näidet sellest protsessist näeb selle saidi alaosas.
Neurotransmitterid. Glutamaati leidub ja see paistab muidu kõige olulisemana, arvestades kui tavaline see on sensoorsetes neuronites ja teisteks olulistemateks neurotransmitteriteks silmanärvi neuronitel on inhibeerivad glütsiin ja GABA. Teiseks ei paista, et leidub teisi aktiveerivaid signaali esilekutsujaid silmas, kuigi inhibeerivat kohalikku kontrolli esineb. Sellega seondumiseks on NMDA ja AMPA retseptoreid.
Silmanärvid ise viivad inimesel signaale ajusse rohkem kui miljoni aksoniga.
 Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala).
Lateraalses genikulaattuumas vahendab nägemissignaale 570 000 neuroniga ja saadetakse signaalid edasi 538 miljonit neuroniga visuaalsele korteksile (Brodmann'i 17. ala). Haistemeelest
Inimesel on 12 miljonit haistenärvi 10 ruutsentimeetrisel pinnal (koeral võib olla 1-4 miljardit lõhna tajuvat neuronit 170 ruutsentimeetril).
Haistenärvid sisaldavad G-valgu seoselisi retseptorvalke, mis aktiveeruvad, kui neile kinnituvad kindlate omadustega molekulid. Tekkinud signaalid kanduvad inimesel aju all olevatesse piklikesse haistesibulatesse (olfactory bulb).
 Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.
Ühe kergelt olulise omadusena võivad sissehingatud aineid selle kaudu ajusse pääseda, kuid mitte väga kergesti. Katses hingasid rotid 12 päeva mangaanoksiidi osakesi (samalaadset tõestatud ka titaanoksiidi ja kulla osakestega) umbes keevitajate töötingimustes oleval tasemel ja eksperimendi lõpus leiti lõhnasibulast 3,5 korda rohkem mangaanoksiidi, kui kontrollgrupis. Kontsentratsiooni tõusu oli ka cerebellumis ja teistes piirkondades. Selles osas on haistenärvil mingit potentsiaali, kuna lõhnaretseptoritega rakud on keskkonnaga suhteliselt otse kontaktis. Peamiseks eelduseks ainete sellisel liikumisel on piisavalt väike suurus ja läbimõõt alla 100 nm paistab ühe olulise omadusena. Vastukaaluks sellele selgitusele kahtlustatakse sissehingamisel muuhulgas hingamisteed selliste osakeste kehas tundlikutesse osadesse transportimisel, kuigi ka seal mainitakse möödaminnes aksonite võimet neid osakesi ajusse viia.Glutamaadi retseptoritest sisaldab lõhnasibul metabotroopseid glutamaadi, AMPA ja NMDA retseptoreid. GABA jaoks on GABA A ja GABA B retseptorid. Aju poolt on reguleeritud atsetüülkoliini, noradrenaliini, dopamiini ja serotoniini eritavate ühenduste poolt. CB1 retseptorid suudavad mõjutada lõhnasibula neuronite aktiivsust.
Maitsmisest
 Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.
Inimesel on kokku 10 000 maitsepunga keelel, suulael ja suu tagaosas. 9000 neist on keelel ja iga maitsepung sisaldab 50-150 retseptorit.Keele maitsepungad sisaldavad G-valgu seoselisi aineid struktuuri järgi ära tundvaid retseptoreid. Aktiveerimisel tekitavad signaali kaaliumiioonide väljavoolu takistamise ja positiivse laenguga ioonide sissevoolu algatamisega. Signaalide maitsepungadest edasi levimine käib ähmasemal viisil, sest rotil näiteks osaleb selles 3 kraniaalnärvi paari. Lisaks 9. kraniaalnärvi paarile on keelega ühenduses lisaks vagaalnärv ja näonärv. Nende maitsele reageerivad osad on kõik sama ajutüve tuumaga ühenduses ja sellest edasi jõuavad need muuhulgas ventraal posterioorse mediaalse taalamuse tuuma kaudu insulani. Selles töös mainiti möödaminnes, et loodi transgeenne hiir, kellel oli magusatajus osalev T1R1 retseptor vahetatud opioidi retseptoriga, misjärel reageerisid nad sellega seonduvat ainet maitstes nagu oleks see magus. Tavahiired nii ei reageerinud. Maitseretseptoreid leidub närilistel suuremal hulgal ka suulaes (isiklikult tundsin suulae tagumise pehmema osaga mee maitset selgelt) ja kurgus ning tugevamamaitselisi toite süües jääb meil järelmaitse tuntavaks palju laiemas piirkonnas, kui ainult keelel.
Signaalide edasikandumises osalevad glutamaadi kainaat, AMPA ja NMDA retseptorid. GABA retseptorid on olemas inhibeeriva rolliga.
Kuulmine
Väliskeskkonnast keskkõrva kandunud õhuvõnkumised põhjustavad kuulmekarvade (mehhanosensoritena toimivad rakud) aktiveerumist. Labürindi seinal olevad vestibulaarsed (tasakaalu) ripsrakud reageerivad 3 eri orientatsioonis olevas poolringi kujulises ruumis oleva vedeliku liikumisele keha liikumise tõttu ja aktiveerivad enda küljes oleva neuroni.
Ajusse viivad auditoorseid signaale umbes 30 000 aksonit. Glutamaadi retseptorid olulised kuulmissignaalide vahendajad. Tasakaaluorgani signaalide vahendajaks glutamaadi AMPA, NMDA ja metabotroopsed retseptorid.
Lapse kuulmisvahemik on 20-20 000 Hz ja vanadel inimestel võib see olla 50-8000 Hz. Ning nende 30 000 neuroni vahendatud signaalid jõuavad 570 000 mediaalse genikulaattuuma neuroni vahendusel auditoorsele korteksile, milles on ~100 miljonit neuronit.
Kuulmiskahjustused tekivad pikajalise ~90 db helitugevuse korral ja valu algab 130 db helide korral.
Lihaskontroll
Tahtlik lihaskontroll toimub alfa mootorneuronite abil ja selleks kasutatakse atsetüülkoliini, mis lihasrakkudel olevate nikotiinsete atsetüülkoliini retseptoritega seondub. Nende aktivatsioon põhjustab naatriumi ioonide sissevoolu ja lihasraku mebraanipotentsiaali muutust sarnaselt neuronitega. Raku siselaendu muutus põhjustab kaltsiumi ioonide järsku vabanemist rakusisestest varudest ning kuigipalju ka rakuvälisest keskkonnast sisse tulekut, mis omakorda algatab mootorvalkude poolset energia kulutamist ja lihasraku kokkutõmmet. Atsetüülkoliin lagundatakse kiiresti või kandub passiivselt rakuvälises keskkonnas mujale. Nikotiinsete atsetüülkoliini antagonistid halvavad tahtlikule kontrollile alluvad lihased ja neid kasutatakse kirurgias patsiendi ajutiseks halvamiseks, sest üldnarkoosis olles võib muidu kerget liikumist ette tulla ja teiseks lõdvestab see lihaseid, lihtsustades ligipääsu lihaste taga olevale.
Oksütotsiinist
Oksütotsiin on vajalik piima eritumiseks ja see on osaliselt neuroloogiliselt reguleeritud. Kui nibu stimuleeritakse puudutusega, siis kanduvad signaalid sellest hüpotaalamusse, mis põhjustab oksütotsiini eritumist ajuripatsist verre. Inimeste puhul võib seetõttu mööduda 30-60 sekundit, enne kui piim hakkab imetamisel erituma. Seal artiklis kirjutati, et kuna aju kontrollib oksütotsiini vabanemist, siis võib selle vabanemine alata ka enne imetamist nagu näiteks nutu kuulmisel.
Mehhano- ja termoretseptorid nahas

Puudutustele ja survele reageerivad mehhanoretseptorid põhjustavad aktsioonipotentsiaali, kui kuju muutus selles olevale neuronile muudab ümbritseva vedeliku survet ja mõjutab ioonide suhet vedeliku sissevoolu mõjutamisega. Tavaliselt on valuretseptoriteks ülaloleval pildil asuvad vabad närvilõpmed, kuid ka need reageerivad puudutustele ja temperatuurile mingil määral.
Temperatuuritajus osalevad temperatuuritundlikud ioonkanalid mis võivad reageerida ka farmakoloogilisele aktiveerimisele. Erinevad temperatuuriretseptorid võivad osaleda neutraalses ja valutekitavalt intentsiivses temperatuuritajus. Mentool aktiveerib külmaretseptorid ja kapsaitsiin tsillipiprast aktiveerib kuumaretseptoreid. Viimane tekitab ka valutaju.
Nervus vagus
N. vagus e. uitnärv või vagaalnärv on kraniaalnärv, mis on nii motoorse kui sensoorse toimega. Teised kraniaalnärvid piirduvad tavaliselt pea ja kaelapiirkonnaga, kuid n. vagus saadab ühendusi jämesooleni (mitte lõpuni). Ühe elutähtsama funktsiooniga saadab see ajust südamesse aeglustavaid ühendusi. Südant stimuleerib närvisüsteem veidi kaudsemalt adrenaliiniretseptorite aktiveerimisega adrenaliini vabastamisega mujalt. Uitnärv vajab atsetüülkoliini oma inhibeerivate signaalide saatmiseks ja seetõttu suudavad atsetüülkoliini blokeerivad ained pulsi väga kiireks ajada. Elustamisel kasutatakse südame liikumasaamiseks või lihtsalt kiirendamiseks adrenaliini või atropiini (atsetüülkoliini blokeerija). Mõlemal on potentsiaali liiga kiire pulsi tekitamisel surmav olla.
Pudendaalnärv
Pudendaalnärv on mõlema soo esindajate genitaalide ja lähiümbruse närv, mis omab sensoorseid ja motoorseid ühendusi. Motoorsed reguleerivad põie ja päraku lihaseid. Sensoorsed pudendaalsed ühendused kannavad paaritumises olulisi signaale sh. mõnu selgroonärvi kaudu ajusse. Signaalid kanduvad edasi peamiselt glutamaadi vahendusel. Selgroonärvist tagasi tulevad ühendused, mis põhjustavad kohalikku verevoolu tõusu ning erutusmuudatusi atsetüülkoliini poolse aktivatsiooniga. Noradrenaliin mõjub erutust inhibeerivana neurotransmitterina. Mingi roll on selliste signaalide kandumisel ka vagaalnärvil.
Selgroonärv
 Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.
Selgroonärvis on umbes miljard neuronit ning tegeleb enamike peast allapoole jäävate lihaste ja meeleelundite signaalide vahendamisega (pilt mõningate ühenduste lähtekohast). Selles on selgelt eristatav kergelt liblika kujuline hall mass ja seda ümbritsev valge mass. Ristlõige on suhteliselt sarnane selja alaosast medullani. Selgroonärviga ühenduses olevate närvide juures on omamoodi perifeersed halli massi kogumid, mis vahendavad keha ja selgroonärvi vahel olevat liiklust. Rakulise ehituse poolest on jagatud kümneks lamina kihiks. Inhibeerijateks on tavaliselt glütsiin ja GABA, mis mõlemad suudavad neuroni siselaengu negatiivsemaks ajamisel raskendada signaali teket.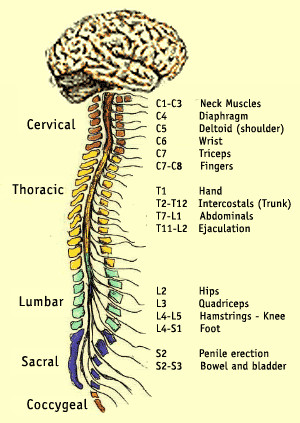{kind=link}
 Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.
Lamina I piirkond (seljapoolsel küljel) vahendab A delta neuronite signaale, mis vahendavad järsku ja teravat valu või kuumatunnet kiirusega 2-30 m/s. Lamina II e. substantia gelatinosa kannab C kiududega aeglaseid, kõrvetavaid valusignaale, sügelust ja puudutust. C kiud on müeliinkihita ja seetõttu näeb see piirkond välja natuke läbipaistvam, kui naaberpiirkonnad ning signaalid liiguvad 0,4-2 m/s. Näiteks näpistusel põhjustavad A delta kiud esialgu tugeva valu ja lõpetamisel saadavad C kiud mõnda aega kõrvetavaid valusignaale. A delta ja C kiud vahendavad signaale vabadelt närvilõpmetelt ajusse ning on perifeerses närvisüsteemis olles kõige väiksema läbimõõduga ja on kõige tavalisemad ühendused perifeersete närvikimpude hulgas. Valusignaalide vahendamisel osalevad substants P (neuropeptiid P) ja glutamaat (NMDA ja teiste retseptoritega). Korduvstimulatsioon tugevdab valu ja selle põhjuseks võib olla vabanenud glutamaadi ja substants P taseme järjestikune tõus. NMDA retseptorid aktiveeruvad pikaajalisema glutamaadi eritumise peale. Inhibeerivatena on opioidi mu ja kappa retseptorid, mida aktiveeritakse periakveduktaalsest hallist massist pärinevate ühendustega. Keha kudedes olevad C kiud suudavad substants P eritamisega aktiveerida histamiini eritavaid valgeliblesid ja laiendada veresooni. Oksütotsiini eritumine hüpotaalamusest nendesse piirkondadesse inhibeerib valusignaale.Nucleus proprius kannab edasi sensoorset infot puudutuste, jäsemete asukohtade, temperatuuri ja valu kohta kohta. Asub lamina III-V piirkonnas. Dorsaalne tuum (lamina VI-VII piirkonnas) vahendab alateadlikke signaale kehaosade asukohtadest cerebellumile ja vigastus põhjustab ataksiat ning kehaosade kordinatsioonihäired. Lisaks vahendavad VI-VII kihid signaale somatosensoorsele korteksile. Signaalid puudutustest ja asukohtadest liiguvad kiirusega 35-120 m/s.
Lamina VII-IX vahendavad motoorseid signaale lihastesse. VII e. intermediolateraaltuum reguleerib lisaks autonoomsetele lihastele veel sümpateetilist närvisüsteemi. Umbes IX piirkonnas asuv Onuf'i tuum saadab isastel rohkem motoorseid ühendusi genitaalidesse kui emastes.
Lamina X ümritseb selgroovedelikku sisaldavat tsentraalkanalit, ühendab selgroonärvi pooli ja vahendab kehast pärinevaid signaale ning reguleerib näiteks põie lihaseid, kuid ülevaatlikumat infot on raske leida. Osaleb parasümpateetilise närvisüsteemi aktiveerimisel ja sealhulgas tegeleb seksuaalses erutuses genitaalide verevoolu regulatsiooniga (mitte erektsioonis).
Lisaks liiguvad signaalid mõlemas suunas ka valges massis (ainult mitte nii korrapäraselt ja funktsionaalselt üleminevalt) ja seal paistab ühe olulisema ühendusena teadvuse jaoks spinoretikulaarne trakt, mis kannab kehast valusignaale ja lõpetab oma ühendused retikulaarses formatsioonis.
2 comments:
Lisaks puudutusele aktiveerub või hakatakse oksütotsiini tootma ka reageerides helidele. Ehk kui imik hakkab nutma, sest tal on kõht tühi, reageerib ema aju valmistades keha ette imetamiseks. Sellisel juhul võib ka piima erituda juba enne puudutust.
Kirjutati küll niimoodi. Esimene kommenteerija, kelle jutust paistab, et on lugenud viitena kasutatud teadustööd :)
Post a Comment