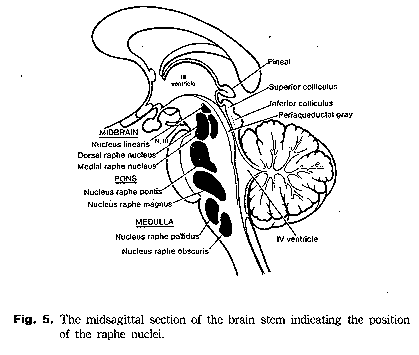
 Raphe (ladina keeles "õmblus") tuumad (RT) on ajutüves lähestikku asetsevad tuumad millest osad on tugevalt stimuleerivad. Need jäävad ajutüve keha külgede vahelisele keskpiirile. Osad keskaju piirkonnas olevad RT'd on kehalist aktiivsust vähendavad ja ponsis olev RT suudab REM une ajal lihashalvatust tekitada. Enamasti eritatakse serotoniini, mis aktiveerib neuroneid ning hüpotaalamusse otsa jõudev serotoniin võib ka palavikku ning unisust tekitada kuna sealsed rakud mõjuvad aktiivselt vastavalt.
Raphe (ladina keeles "õmblus") tuumad (RT) on ajutüves lähestikku asetsevad tuumad millest osad on tugevalt stimuleerivad. Need jäävad ajutüve keha külgede vahelisele keskpiirile. Osad keskaju piirkonnas olevad RT'd on kehalist aktiivsust vähendavad ja ponsis olev RT suudab REM une ajal lihashalvatust tekitada. Enamasti eritatakse serotoniini, mis aktiveerib neuroneid ning hüpotaalamusse otsa jõudev serotoniin võib ka palavikku ning unisust tekitada kuna sealsed rakud mõjuvad aktiivselt vastavalt.Tavaliselt inhibeeriva mõjuga GABA agonistide süstimisel mediaalsesse ja dorsaalsesse raphe tuuma ning ventraalsesse tegmentaalsesse alasse põhjustas hüperaktiivsust ning suurendas tarbitud toidu ja joogi hulka. Kaasnes suurem dopamiini tarbimine (tõenäoliselt suurema dopamiini vabanemise tagajärg) striaatumis ja nucleus accumbens'is kuid serotoniini hulk striaatumis ja hippokampuses püsis samal tasemel.
Võib-olla nende keskaju Raphe tuumade pärast ongi inhibeerivad GABA agonisti tüüpi rahustid tegutsemistahet ja heaolutunnet andvad.
Mediaalse RT ja selle lähiümbruse rakkudel on korduvalt leitud liikumist inhibeerivat toimet. Glutamaadi agonisti süstimisel vähendas see kohalike rakkude aktiveerimise järgselt katseloomade liikuvust.
Mediaalne RT on üks suuremaid signaali saatjaid septumile, amügdalale ja hippokampusele. Sellest pärinev serotoniin on ehmatuse ajal üks mälualade stimuleerijaid.
Immuunsüsteemi aktiveerimisel vähetoksiliste ja surnud bakteritega suurenes aktiivsus ajutüves rohkem dorsaalses RT's. Palavikureaktsioonid on suuresti serotoniini poolt läbi viidud. Ka osad serotoniini vabastavad ained võivad liigse palavikuga äärmisel juhul surmavalt mõjuda. Suurem serotoniini tarbimine paistis nakkuse ajal frontaalsagara alaosas.
Enamus serotoniini eritavaid neuronid on aktiivsed ärkvel olles ja vaigistuvad REM une ajal. Erandina paistab dorsaalse ja mediaalse RT alaosas neuroneid, mis aktiveeruvad sõltumata ärkvelolekust palaviku ajal. Seejuures võib limbilises süsteemis tõusta serotoniini vabanemine kuigi emotsioonid nõrgenevad ja tekib depressiivsus.
Dorsaalne RT on aktiivsem negatiivsete depressiivsete ja stressis tujude ajal nagu näiteks vangistuse või õpitud abituse ajal allaandlikult kannatamisel. Ühendusi on ärevuses osalevate amügdalaga ja selle naabertuumadega.
Dorsaalne RT on üks valusignaalide takistaja läbi suure RT (pildil nucleus raphe magnus) minevate ühendustega. Dorsaalse RT enkefaliine ja serotoniini sisaldavad rakud on valu vähendavad ja selle GABA sisaldavad rakud on valureaktsioone tugevdavad.
Agrüpnia korral esineb täielik unetus ilma väsimustundeta. See võib tekkida RT, taalamuse ja otsmikupoolse hüpotaalamuse kahjustuse järgselt. Surmavat unetust tekitava fatal familiar insomnia puhul kahjustub kõige põhjalikumalt taalamust.
Dorsaalne RT osaleb ärkveloleku alahoius stimuleerides prefrontaalset korteksit. Umbes 75% ühendustest sisaldavad serotoniini. Dorsaalne RT (DRT) ise saab hüpotaalamusest oreksiini. Oreksiini süstimine DRT sisse suurendab selle aktiivsust (rottidel üle 10 minuti) ning muutus on suurem sügava või REM une ajal süstides. DRT aktiivsus on kõrgeim ärkvel olles ning REM une tekkeks peab selle tegevus minimaalseks minema. Autorite järgi võivad narkoleptikute raskused ärkvelolekus olla tingitud oreksiini puudusest muuhulgas DRT's.
Retikulaarne formatsioon
 Retikulaarne võrgustik on peamiselt ajutüves asuv võrestikulaadne struktuur, mis ühendab peaaegu kõiki sel asuvaid tuumi ning selle ühendused ulatuvad ajutüvest välja taalamusele, ajukoorele ja selgroonärvi alaosani. See kontrollib paljusid ehmatusreaktsioone saates 10-120 meetrit sekundis signaale selja, kaela, õlgade ja jäsemete torsopoolsete alguskohtade lihaste motoorsetele neuronitele selgroonärvi hallis massis põhjustades ehmumisel nende aktiivsemaks minekut.
Retikulaarne võrgustik on peamiselt ajutüves asuv võrestikulaadne struktuur, mis ühendab peaaegu kõiki sel asuvaid tuumi ning selle ühendused ulatuvad ajutüvest välja taalamusele, ajukoorele ja selgroonärvi alaosani. See kontrollib paljusid ehmatusreaktsioone saates 10-120 meetrit sekundis signaale selja, kaela, õlgade ja jäsemete torsopoolsete alguskohtade lihaste motoorsetele neuronitele selgroonärvi hallis massis põhjustades ehmumisel nende aktiivsemaks minekut. Pildil on näha medulla retikulaarne formatsioon ning sellele omane ühtlane võrgustiku laadne rakkude paigutus, mis on nimetuse (ladina keelest võrgu laadne) põhjus.
Pildil on näha medulla retikulaarne formatsioon ning sellele omane ühtlane võrgustiku laadne rakkude paigutus, mis on nimetuse (ladina keelest võrgu laadne) põhjus.
Illustratsioon suurema ponsi alaosas asuva RF neuroni ühendustest. Sellest algav akson hargneb T kujuliselt kaheks ja võib ulatuda ühes suunas taalamuse ümbrusesse, sisemusse ja ajukooreni ning teises suunas selgroonärvi alaosani. Ühendab ka näo, kõri, keele ja silmade sensoorsete ja motoorsete kraniaalnärvide tuumasid. Sellised suured rakud moodustavad ~5% RF rakkudest. Taalamuse ümber olevat retikulaarset tuuma ei loeta RF osaks ja selle puhul on ühendused pärit kohalikest rakukehadest.
 Kuigi üheks algseks nimetuseks oli retikulaarne aktivatsiooni süsteem, sest see võib erksust põhjustada, ei paista ajutüves ega ajus ärkvel olekuks rangelt vajalikke osasid kuid une tekkeks vajalikke osasid esineb RF's.
Kuigi üheks algseks nimetuseks oli retikulaarne aktivatsiooni süsteem, sest see võib erksust põhjustada, ei paista ajutüves ega ajus ärkvel olekuks rangelt vajalikke osasid kuid une tekkeks vajalikke osasid esineb RF's. Retikulaarne formatsioon (RF) saab signaale kõikidelt sensoorsetel ja motoorsetelt neuronigruppidelt, mis ajusse jõuavad või sellest väljuvad. RF reguleerib neile signaalidele tuginedes ülejäänud närvisüsteemi aktiivsustaset olles osaliselt harjumise läbiviija. Selle osana ei arvesta sinna piirkonda jäävaid kraniaalnärvi tuumasid, medulla oliivi ja substantia nigra't. Kuna suur osa RF'st põhjustab stimuleerimisel aeglaste ajulainete sünkroonsuse kadumist nimetati seda piirkonda retikulaarseks aktivatsioonisüsteemiks, kuigi kõik selle osad pole stimuleeerivad ja see nimi on ülelihtsustav.
Retikulaarne formatsioon (RF) saab signaale kõikidelt sensoorsetel ja motoorsetelt neuronigruppidelt, mis ajusse jõuavad või sellest väljuvad. RF reguleerib neile signaalidele tuginedes ülejäänud närvisüsteemi aktiivsustaset olles osaliselt harjumise läbiviija. Selle osana ei arvesta sinna piirkonda jäävaid kraniaalnärvi tuumasid, medulla oliivi ja substantia nigra't. Kuna suur osa RF'st põhjustab stimuleerimisel aeglaste ajulainete sünkroonsuse kadumist nimetati seda piirkonda retikulaarseks aktivatsioonisüsteemiks, kuigi kõik selle osad pole stimuleeerivad ja see nimi on ülelihtsustav.Osad RF rakud osalevad valutaju reguleerimises saates aksoneid selgroonärvi dorsaalsetele sarvedele.
RF sisse jäävad aju suuremad noradrenaliini, atsetüülkoliini, dopamiini ja serotoniini eritajad. Locus coeruleus'i tuumad ponsis suudavad inimese närvisüsteemi noradrenaliiniga varustada kahe 30 tuhande rakulise tuumaga mõlemal kehapoolel. Noradrenaliin on lühiajaliselt tugeva stimuleeriva toimega, kuid locus coeruleus'i rakud aeglustavad tegevust pikemaajalise stiimuli korral ja atsetüülkoliinirakkude aktiivsus suureneb tähelepanu säilitamiseks.
RF saab signaale tihedamalt pea piirkonnast ja kõige vähem torsost ning alajäsemetest. Tagajäseme närvi tihedal stimulatsioonil võib neuronite töösagedus langeda minutiga 10 korda erinevalt visuaalse korteksi neuronitest, mille aktiivsus langeb kuni 10-15% ning selle erinevuse põhjusena paistab RF abil kohanemine.
Medulla mediaalse RF stimulatsioon on motoorseid neuroneid inhibeeriva toimega.
Kuigi RF on järjepidev piirkond läbi ajutüve leidub selles spetsiifilisemalt kindlatele kehaosadele mõjuvaid alasid. Medulla alaosasse jäävad alad osalevad rohkem kaela ja selja lihaste kontrollis, kuid kõrgemale ja ajutüve keskosale lähemale jäävad osad mõjuvad järjest hajusamalt ja laiemale jäävaid lihaseid.
Rf ülaosas hargnevad selle ühendused keskaju järel kahes suuremas suunas. Üks haru läheb taalamusele. Teine haru läheb hüpotaalamusele, septaalalale ja umbes 10% jõuab ajukooreni (kõige rohkem ajukoore otsmikupoolse kolmandikuni).
Taalamuseni ulatuvat haru paistab sünkroonsete ajulainete nagu näiteks une alguses olevate sünkroonsete kiirete signaalide (unespindlite) ja puhkeolekus olevate ~10 Hz ajulainete tekitajana.
Ajukoorele ja septumile minev haru paistab sünkroonsete ajulainete kaotajana nii magades kui ärkvel olles. Ärkvel olles võib see ajukoort stimuleerida ning une ajal on see oluline REM une ajal sünkroonsuse kaotamisel. Retikulaarsest formatsioonis olevatelt Raphe tuumadelt algavad ühendused ulatuvad septaalale ja on hippokampuses aktiivsemal ajal olevate ~8 Hz teeta lainete algatajad.
Taalamuse sisse jäävad seda läbivad lehtja tuumad, mis osalevad ajulainete tekitamises.
 Pildil paistab selliste osade hulka jääv valgelt tähistatud piirkond koos tsentromediaalse ja interlaminaarse tuumaga.
Pildil paistab selliste osade hulka jääv valgelt tähistatud piirkond koos tsentromediaalse ja interlaminaarse tuumaga. Lisaks paistab ühe tekkepõhjusena ajulainete tekkele taalamusi läbiv ja ühendav internal medullary lamina, mis paistab kergelt liblika kujulisena.
Lisaks paistab ühe tekkepõhjusena ajulainete tekkele taalamusi läbiv ja ühendav internal medullary lamina, mis paistab kergelt liblika kujulisena.Sellisel läbival tuumal paistab potentsiaali teiste taalamuse piirkondade mõjutamises oma tegevusega, sest kui selles peaks tekkima samaaegselt tegevus võivad kõik teised taalamuse tuumad saada tugeva signaali mis piisava tugevuse korral ka nendest ja nendega ühendatud aladest läbi käiks. Magamise ajal kaasnenud langus dopamiinis, serotoniinis ja adrenaliinis aitaks ülejäänud taalamusel vaigistuda ja lasta atsetüülkoliiniga aktiveerunud ajulaine tekitajatel üle kogu taalamuse samaaegseid signaale väljastada.
 Elektroodidega ajukoorelt mõõdetud tegevus on REM une ja ärkveloleku ajal väga sarnased, kuid sügava unenägudeta une ajal esineb suuri elektrilisi kõikumisi, mis tähendab rohkemate neuronite samaaegset aktiveerumist. Teadvusetus olekus on sellised samaaegsed (sünkroonsed) lained kõige selgemalt näha.
Elektroodidega ajukoorelt mõõdetud tegevus on REM une ja ärkveloleku ajal väga sarnased, kuid sügava unenägudeta une ajal esineb suuri elektrilisi kõikumisi, mis tähendab rohkemate neuronite samaaegset aktiveerumist. Teadvusetus olekus on sellised samaaegsed (sünkroonsed) lained kõige selgemalt näha.Keskaju RF tegevust mõõdeti valgussähvatuste ja heliklõpsude ajal. Valgus tekitas suuremat aktiivsust kuklapoolsel keskosal ning helid rohkem külgedel ja näo pool.
Ponsi alaosa retikulaarse formatsiooni suurerakulise piirkonna (pildil suuremate laikudega) kahjustamisel nõrgenes või kadus tugeva heli kuulmisel ehmatusreaktsioon. Need rakud saavad signaale helitajus osalevatelt aladelt ja saadavad signaale selgroonärvi motoorstele neuronitele.
Parvocellular RF oli vahendusteeks medulla NTS'st ponsi parabrahhiaaltuumadeni. NTS ülaosa saab maitsesignaale ja alaosa vagaalnärvi signaale, kuid need signaalid ei aktiveeri selles samu piirkondi. Selle RF kaudu jõuavad signaalid parabrahhiaaltuumadeni (ja tagasi), kus signaalid jõuavad mõlema NTS osa stimuleerimiskatsete järgi samadele rakkudele. Autorite järgi võisid need ühendused ala hoida ajuta katseloomade maitseelistusi ja muutusi isus kõhu täitumisel.
Süstides värvi erinevatesse parvocellular (suuremarakulistesse) RF piirkondadesse värvusid selle ülaosa puhul tihedalt kolmiknärvi keskajusse jääv osa koos viimase kohal oleva alaga. Tihedamalt värvusid veel näonärvi ja keelaluse närvi tuumad koos parabrahhiaaltuumadega. Keskmiselt tihedusega oldi ühenduses kolmiknärvi spinaalse tuumaga, NTS'ga, medulla RF, oliivi ja motoorsete RF aladega. Ühendusi polnud vagaalnärvi motoorse tuumaga.
RF'st selgroosse minevad ehk retikulospinaalsed aksonid mõjutavad tugevalt kaela, torso ja jäsemete alguskohtade lihaseid. Tavaliselt jõuavad aksonid mõlema selgroonärvi külje premotoorsetele neuronitele, mis omakorda on ühenduses motoorsete neuroniga. Akson võib sõltumata päritolu kehapoolest haruneda oma otsas mõlemale selgroonärvi küljele. Kui ajukoorelt või punastelt tuumadelt alanud neuronid kontrollivad rohkem jäsemete otsi, siis retikulospinaalsed ühendused soosivad kogu keha lihaste aktiveerumist olles mõjukamad selgroo, puusade ja õlgade piirkonnas. Ajukoorelt tuleb signaale rohkem planeerimisega seostatud premotoorselt korteksil ja vähem primaarselt motoorselt korteksilt. Suurim tihedus paistab nende kahe vahelisel üleminekualal. Esijäsemete ajukoorelt võib veidi rohkem ühendusi alata. Teisteks suurimateks RF'le signaalide saatjateks on cerebellum, vestibulaarsed tuumad, nelikkünkad ning somatosensoorsed ja keha asukohataju signaalid. Kergel naha silitamisel võib RF rakkudes reaktsioon puududa kuid liigesepiirkonna kõõlused on tugevad RF mõjutajad nagu ka valu.
Signaalide levik ajutüvest selgroogu on 10-120 meetrit sekundis.
Motoorse mõjuga RF rakkude hulk suureneb ajutüve alaosas ja ponsi RF stimulatsioonil tekib tavaliselt pea pöördumist samale küljele, sama külje esijäseme kõverdumist ja sama külje tagajäseme sirutumist.
Rütmiliste liikumisliigutuste algatajana on RF paistnud ühe kõige olulisema struktuurina saades signaale keskajus ja cerebellum'is asuvatelt liikumise algatajatelt. Olulisteks on seejuures ka kehast tulevad somatosensoorsed signaalid.
Kassi ponsi-medulla RF'l asub esijäsemetele vastav ala rohkem kõrguselt keskel kukla pool ja tagajäse selle üla- ja alaosa näopoolsel osal. Kaelalihaseid kontrollivad alad asuva sellel kogupikkusel.
Primaatidel põhjustas RF ja tasakaaluelunditest selgroogu minevate ühenduste kahjustamisel raskusi kehaasendi säilitamisel ja peaaegu suutmatus jäsemeid kehast eemale liigutada lihasnõrkuse tõttu. Käelabade kontroll säilis paremini, kuid vahepealsete käelihaste nõrkus tegi nende kasutamise raskeks.
 Pildil on näha kaks peamist RF päritoluga ühendusraja aktiivsused ärkvel olles, sügava une ajal ja REM une ajal. Aminergic tähistab noradrenaliini, serotoniini ja dopamiini ning cholinergic atsetüülkoliini. Mustade täppide tihedus näitab vastavate neurotransmitterite vabanemist sellel ajal.
Pildil on näha kaks peamist RF päritoluga ühendusraja aktiivsused ärkvel olles, sügava une ajal ja REM une ajal. Aminergic tähistab noradrenaliini, serotoniini ja dopamiini ning cholinergic atsetüülkoliini. Mustade täppide tihedus näitab vastavate neurotransmitterite vabanemist sellel ajal.Keskaju RF saadab palju ühendusi taalamusele põhjustades sünkroonsete ajulainete kadumist REM unes ja ärkvel olles.
Keskaju-ponsi piirilt pärit atsetüülkoliin läheb osaliselt taalamusele inhiberides magades aktiivset taalamuse retikulaarset tuuma ja aktiveerides ärkvel olekuks vajalikke rakke.
Hüpotaalamusele ja ajukoorele serotoniini eritava anterioorse/superioorse RT kahjustus võib kaotada unisuse ja une päevadeks. See on aktiivne ärkvel olles ning olulised ühendused on otsmikupoolse hüpotaalamusega. Kahtlustati, et see mõõdab osaliselt ärkvelolekut ja inhibeerib ennast hüpotaalamusega toimuva signaalivahetuse tõttu.
Ükski neist ei paista ärkvel olekuks tingimata vajalikuna ja kahjustuse või farmakoloogilise mõjutamise järel taastub tavaline ärkvel olek mõne päevaga.
 Pildil on illustreeritud REM und algatav RF osa ponsis. Locus coeruleus inhibeeritakse sellel ajal kõige aeglasemaks üldse une-ärkveloleku tsüklis. Atsetüülkoliin taalamusse vähendab aeglaste ajulainete osakaalu ja selgroonärvis lõdvestatakse lihased GABA sarnase inibeeriva toimega glütsiini vahendusel. REM unele nime andnud silmade kiirete liigutuste põhjusena paistab ponsi RF'st keskaju ülaküngaste motoorsetele sügavatele kihtidele minevad ühendused, mis omakorda saadavad signaale silmaliigutusi ajastavale ja suunavale alale ponsi RF's (paramediaalne ponsi RF).
Pildil on illustreeritud REM und algatav RF osa ponsis. Locus coeruleus inhibeeritakse sellel ajal kõige aeglasemaks üldse une-ärkveloleku tsüklis. Atsetüülkoliin taalamusse vähendab aeglaste ajulainete osakaalu ja selgroonärvis lõdvestatakse lihased GABA sarnase inibeeriva toimega glütsiini vahendusel. REM unele nime andnud silmade kiirete liigutuste põhjusena paistab ponsi RF'st keskaju ülaküngaste motoorsetele sügavatele kihtidele minevad ühendused, mis omakorda saadavad signaale silmaliigutusi ajastavale ja suunavale alale ponsi RF's (paramediaalne ponsi RF).REM unega kaasneb peaaegu täielik lihashalvatus vaatamata ajukoore suurele aktiivsusele ka primaarsel motoorsel korteksil. Vananedes võib see lihaste lõtvus väheneda ning une ajal võidakse rohkem liigutama hakata. Motoorsetest signaalidest pääsevad minimaalse takistusega lihasteni silma, keskkõrva ja kopsulihaste lihaste signaalid kuigi hingamisteede lõtvumine võib norskamist tekitada. REM une ajal ponsis vabanev atsetüülkoliin aktiveerib suuri medulla RF rakke, mis eritavad selgroonärvi glütsiini.
REM une ajal tekkivad ponsist läbi lateraalse genikulaattuuma kuklasagarale jõudvad ajulained (PGO lained) algavad samuti atsetüülkoliini mõjul locus coeruleus'ist allapoole jäävatest jäävast piirkonnast. Serotoniin noradrenaliiniga takistab ponsis PGO lainete ja teiste REM und tekitavate rakkude tegevust, kuid sügava une faasist REM une faasi minekul väheneb nende vabanemine piisavalt inhibeeriva mõju kaotuseks.
REM unega kaasneb tavaliselt ka erektsioon mõlema soo esindajatel.
Kuutõbisuse korral oldakse tavaliselt sügava une faasis ning äratamisel ei mäletata unenägusid. REM unele sarnast lihashalvatust esineb narkoleptikutel katapleksia nime all (põhjusena paistab hüpotaalamusest pärit oreksiini puudus) ja ka unehalvatuse korral esineb sellist lihaslõtvust isegi kui teadvus säilib. Narkolepsiahoo ajal magama jäämisel toimub järsk REM une algus, mis võib ka püstiseisjatel tekkida.
Nende häirete korral peaks ka lihaslõtvust vahendama medulla suured RF rakud, kuid tõenäoliselt teistelt rakkudelt tulnud signaalidega, kui REM une ajal. Vananedes surevad lihaslõtvust tekitavad rakud osadel (rohkem meestel) nii palju, et hakatakse REM une faasis unenägusid välja elama.
RF sai oma teise nime (retikulaarne aktivatsiooni süsteem) seoses sellega, et selle kahjustus võis põhjustada pikaajalist und või koomat ning une ajal stimuleerimisega kaasnes ärkamine. Selle ülelihtustatud nimetuse kasutamist vähendati kuna paari kahjustuse järgse päevaga taastus ärkvel veedetud aeg ning osad kahjustused vähendasid magamisele kuluvat aega.
Une tekkeks piisas sobivast taalamuse stimulatsioonist ning sensoorsete signaalide blokeerimine ajust vigastustega ei muutnud une-ärkveoleku tsükli kestvust. REM une ajal on muidu primaarne visuaalne korteks väheaktiivne, kuid keerulisemaid pilte ja stseene vahendavad kuklast veidi kaugemale jäävad visuaalsed alad aktiveeruvad tugevamalt. Primaarse visuaalse korteksi naaberalade stimulatsioonil võib stimulatsioonil tekkida stseenide ja objektide nägemist. Algus REM unele algab siiski ponsi kaudu.
Kõrvalisem hüpotees retikulaarse formatsiooni mõjust näoilmetele ja emotsioonide väljendusele
Selle teema kohta lugedes hakkas paistma rohkem loogikat näoilmete osas. RF osaleb üldiselt närvisüsteemi aktiveerimises intentsiivsemate tajude korral ning selle juurde käivad ka muutused näoilmetes, mis võivad sarnaneda teiste imetajate näoilmetega. See hüpotees tundus väga lihtsa ja veakindlana pisikeste arenguliste kõrvalekallete suhtes proportsioonide ja neuronite töösageduste muutustele.
Tavaline kerge ehmumisreaktsioon või ka teised emotsionaalsed perioodid põhjustavad silmade suuremat avamist ja hajusalt ka teiste näo lihaste kokkutõmmet. Suurema ehmatuse korral millegi tulemisel näo juurde toimub ka silmade ja suu kinni minek. RF paistab hajusa ühendajana kõigile ajutüve tuumadele kuid seejuures ei paista täiesti ühtlaseid ühendusi ning lähinaabrid kalduvad üksteisele suuremat mõju avaldama. Näo ja kõri piirkonna kraniaalnärvide artiklis olid näo, suu ja kõri motoorsed ning sensoorsed tuumad üksteisega ühenduses. Ühe näopoole motoorsed ja sensoorsed aksonid jõuavad samale ajutüve külje või mõlemale külje tuumadele.
Neutraalse erksuse korral võib suu püsida horisontaalselt sirgena kuid positiivsete ja negatiivsete emotsionide puhul on näolihastel tuntud kalduvus vastavalt ülespoole või allapoole tõmbuda.
Vastikustunde korral tõmbuvad rohkem kokku näo alaosa ja kõri lihased, ms tõmbavad veidi kaasa ka näolihaseid. Medullasse jäävad selle piirkonna tuumad koos oksendamist põhjustava vagaalnärvi motoorse tuumaga. Ka tugevad maitsesignaalid nagu hapu ja mõru võivad näo alaosa rohkem pingesse ajada saates signaale medullas olevasse NTS'i.
Halbade lõhnade ja vastikustunnet tekitavate olukordade puhul tõmbuvad näo lihased nina suunas kokku.
Kaudsema ebameeldivustundena tõmbuvad ereda valgusega silmade lähiümbruse lihased kokku.
Positiivse tuju korral tõmbuvad lihased näo ülaosa suunas tõmmates lihaseid silmade suunas. Silmaümbruse lihased tõmbuvad kokku liigutades põsesarnasid silmade suunas ning ka suu ümbrus tõuseb kõrgemale näidates hambaid. Positiivseid elamusi on inimestel ja näiliselt ka katseloomadel saadud keskaju piirkonnas substantia nigra, sealse tegmentumi ja osaliselt keskajusse ulatuvate subtalaamsete tuumade stimulatsioonil. Sinna piirkonda jäävad ka 2 silmalihaste tuuma ja lõualihaseid liigutav kolmiknärvi osa mis võib RF aktiveerumisel põhjustada suu avamist rõõmuga. Heaolutunne tundub mõnutunne vähemalt isiklikult kusagil silmade kohal või nende juures asuvana. Hea tujuga ja silmad kinni on see selgem kuigi asukohatunne on selliste emotsioonide puhul häguse asukohaga.
Naer ise võib tekkida sellest, et RF ajab tugeva serotoniini ja dopamiini poolse hajusa stimulatsiooniga näo, kurgu, kõri ja kopsulihaste lihased pingesse ning nagu kõigile keha lihastele omane kaasneb tugevale venitusele kiiresti vastupidine liigutus ning naeruhoo ajal jõuavad lihased ~5 korda sekundis aktiveeruda ja lõdvestuda. Naeru ajal toimuvat kiiret ja põhjalikku välja hingamist saavad põhjustada ponsi parabrahhiaaltuumad. Sissehingamiseks vajalik jääb medullasse ning ponsi ja medulla vaheline konkurents kopsude üle võib naeruhäälitsusi algatada. Isiklikult on meeldiva enesetunde korral kergem ja mõnusam välja hingamine. Ärevusega on kergem ja kiirem sisse hingamine. Nutu korral võivad intentsiivsed emotsioonid samuti analoogselt lühikesi rütmilisi lihaskokkutõmbeid põhjustada. RF laialdased ühendused võivad nutu ja naeruga põhjustada veel pisaranäärmete aktiveerumist koos nina-kurgu-kõri piirkonna limanäärmete aktiveerimisega. RF vahendatud hajusate ühenduste võrestik võibki olla põhjuseks sellele miks kindlat naeru või nutu tuuma pole leitud kuigi juba 1924. aastal suutis Kinnear Wilson piiritleda need kaks ponsi piirkonda. Patoloogiline emotsioonitu naer ja nutt võivad siis ka alata näo ja kõri piirkonna rakkude ebanormaalsel tegevusel. Esimese hooga võib kohalik hüperaktiivsus tekkida sellega, et surevad rakud lähevad lõpus hüperaktiivseks suutmata energiapuuduse tõttu siselaengut normaliseerida ning hilisema tekkega naer võib tekkida sisendita (motoorsed ühendused ajukoorelt) jäänud neuronikogumitest mis analoogselt spastilisuse või neuropaatilise valuga hakkavad iseseisvalt impullse algatama mõjudes naeru tekitavalt.
No comments:
Post a Comment